Diversity, Bioactivity Profiling and Untargeted Metabolomics of the Cultivable Gut Microbiota of Ciona intestinalis

 , , and
, , and
Abstract
:
1. Introduction
2. Results
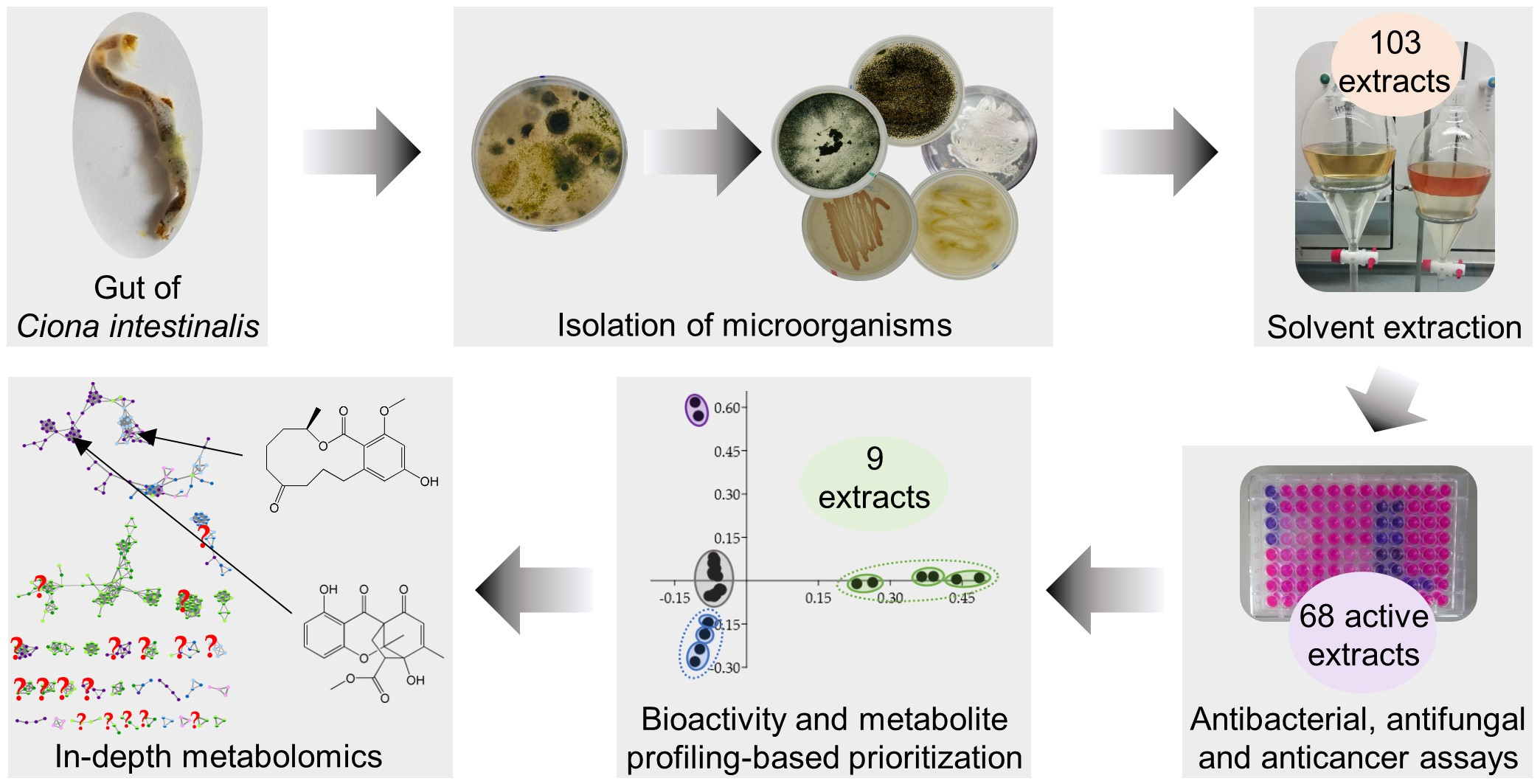
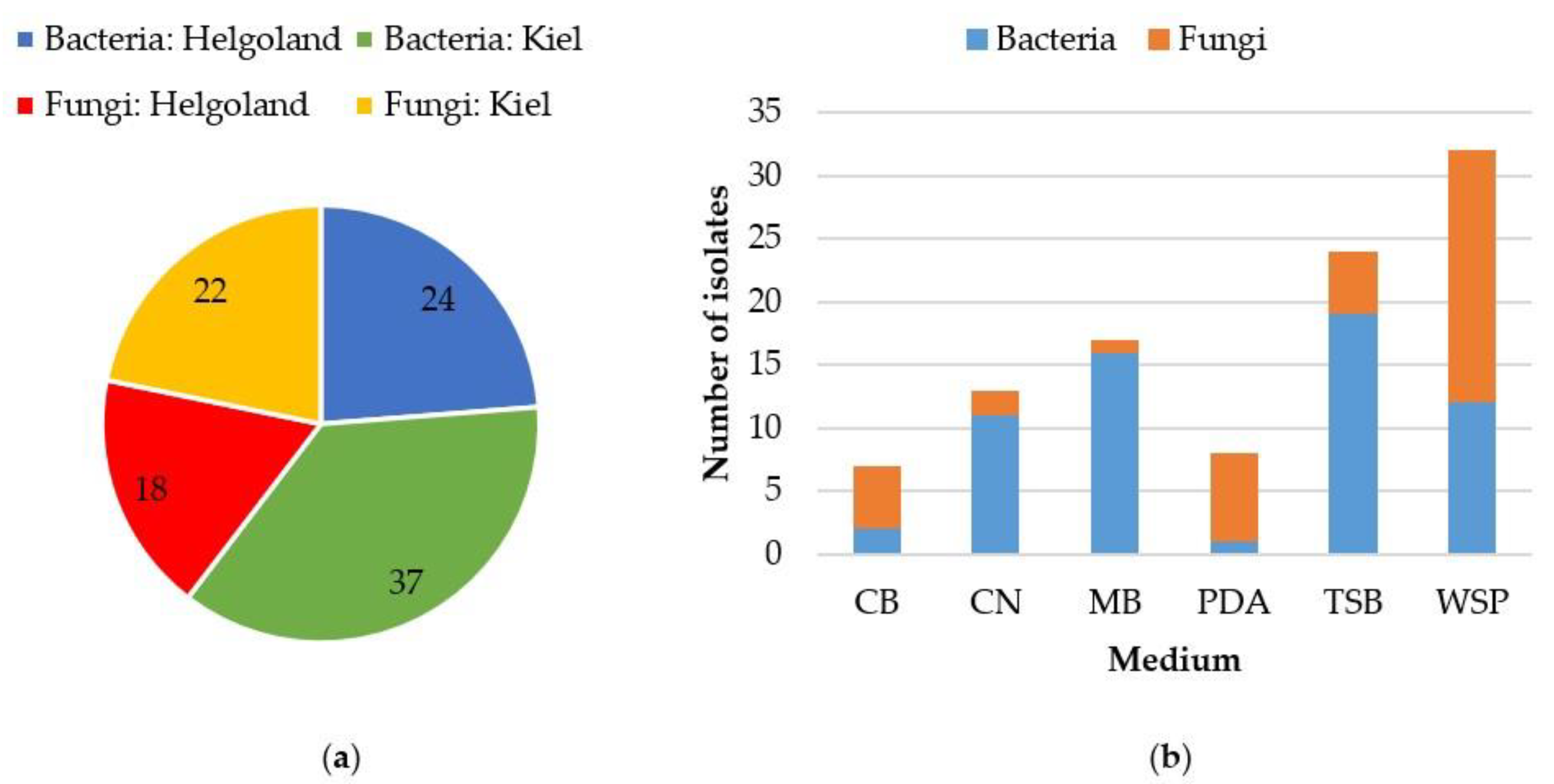
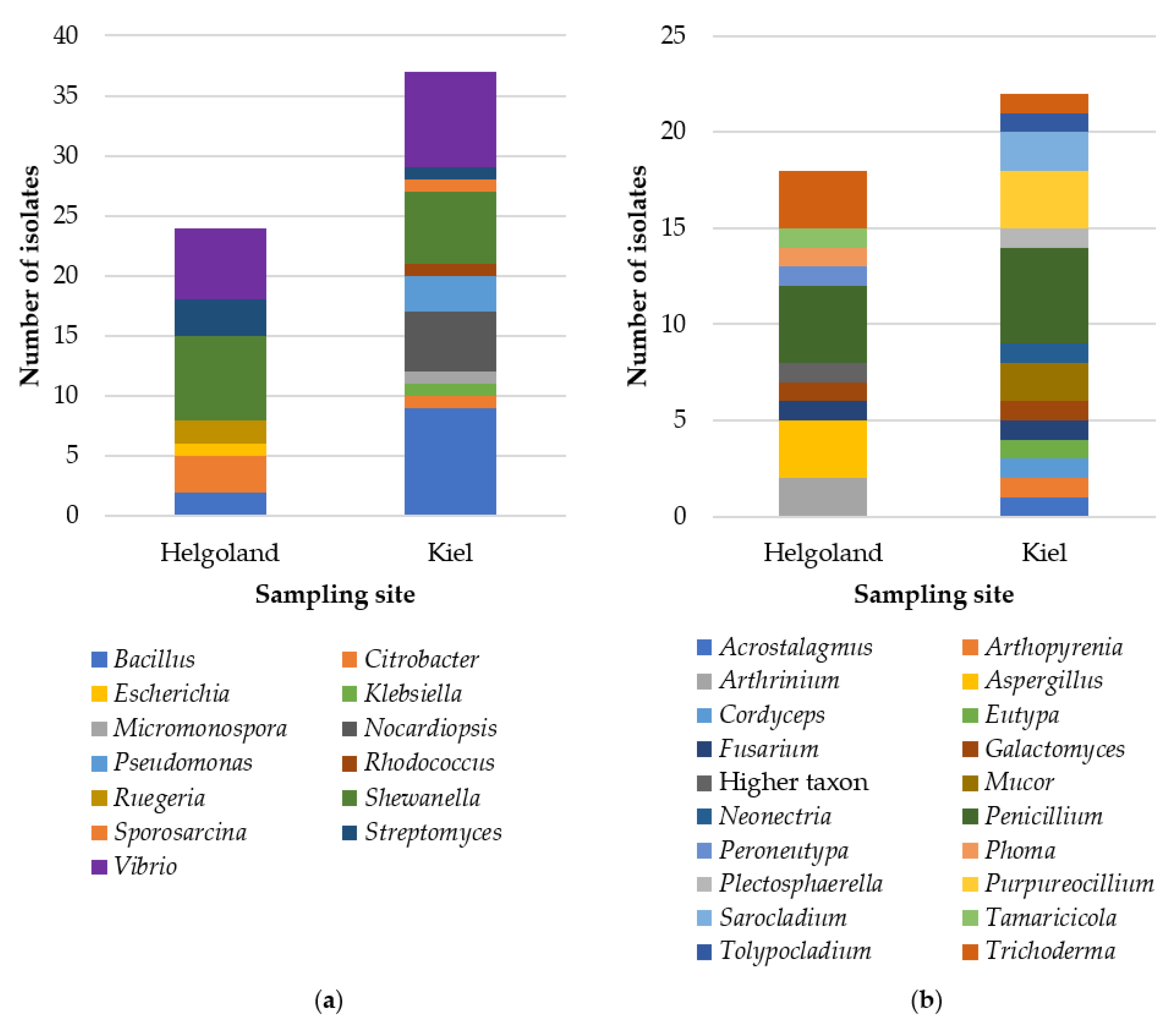
2.1. Cultivable Fraction of the Gut Microbiota of C. intestinalis
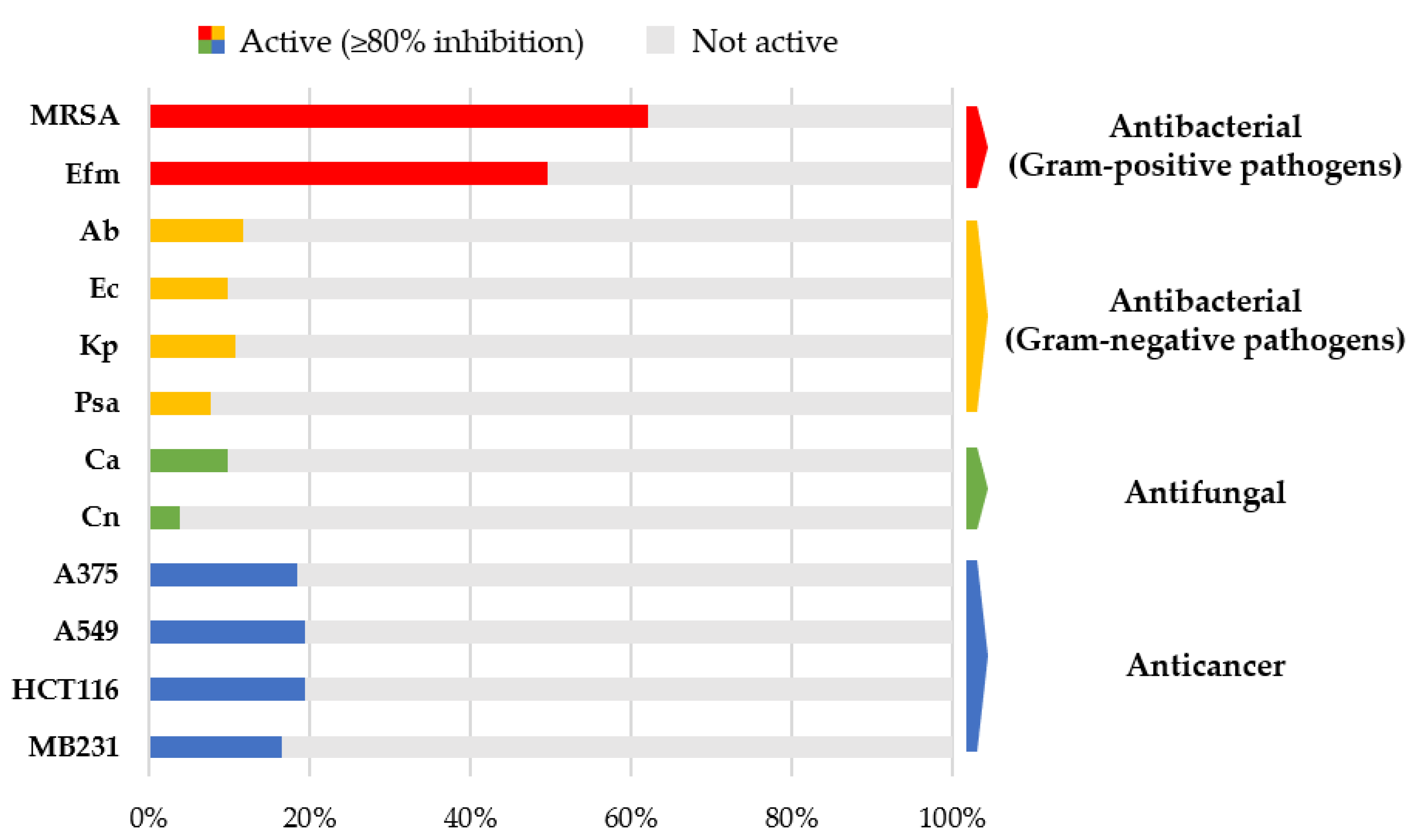
2.2. Biological Activities of Gut-Derived Microbial Extracts
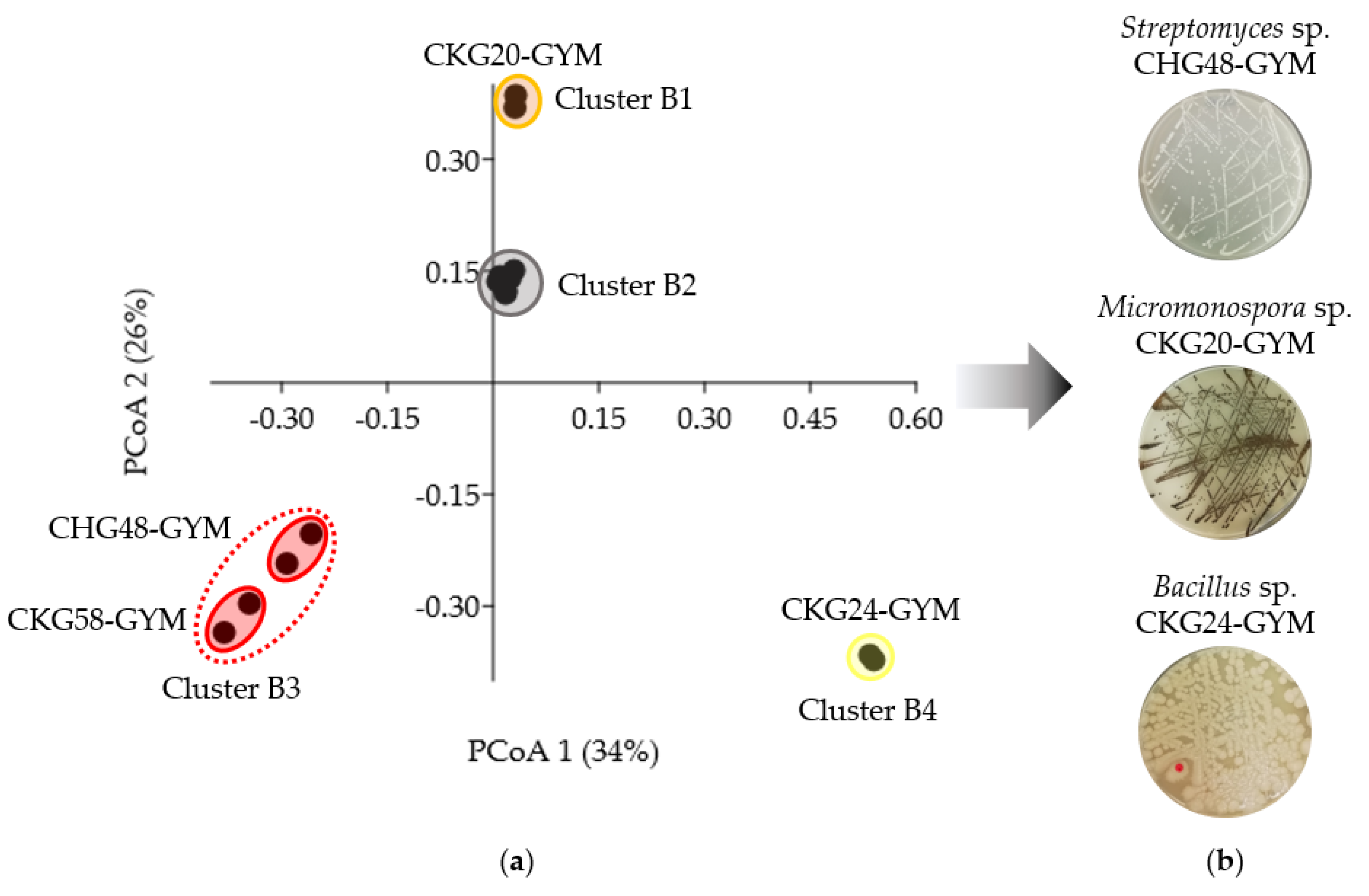
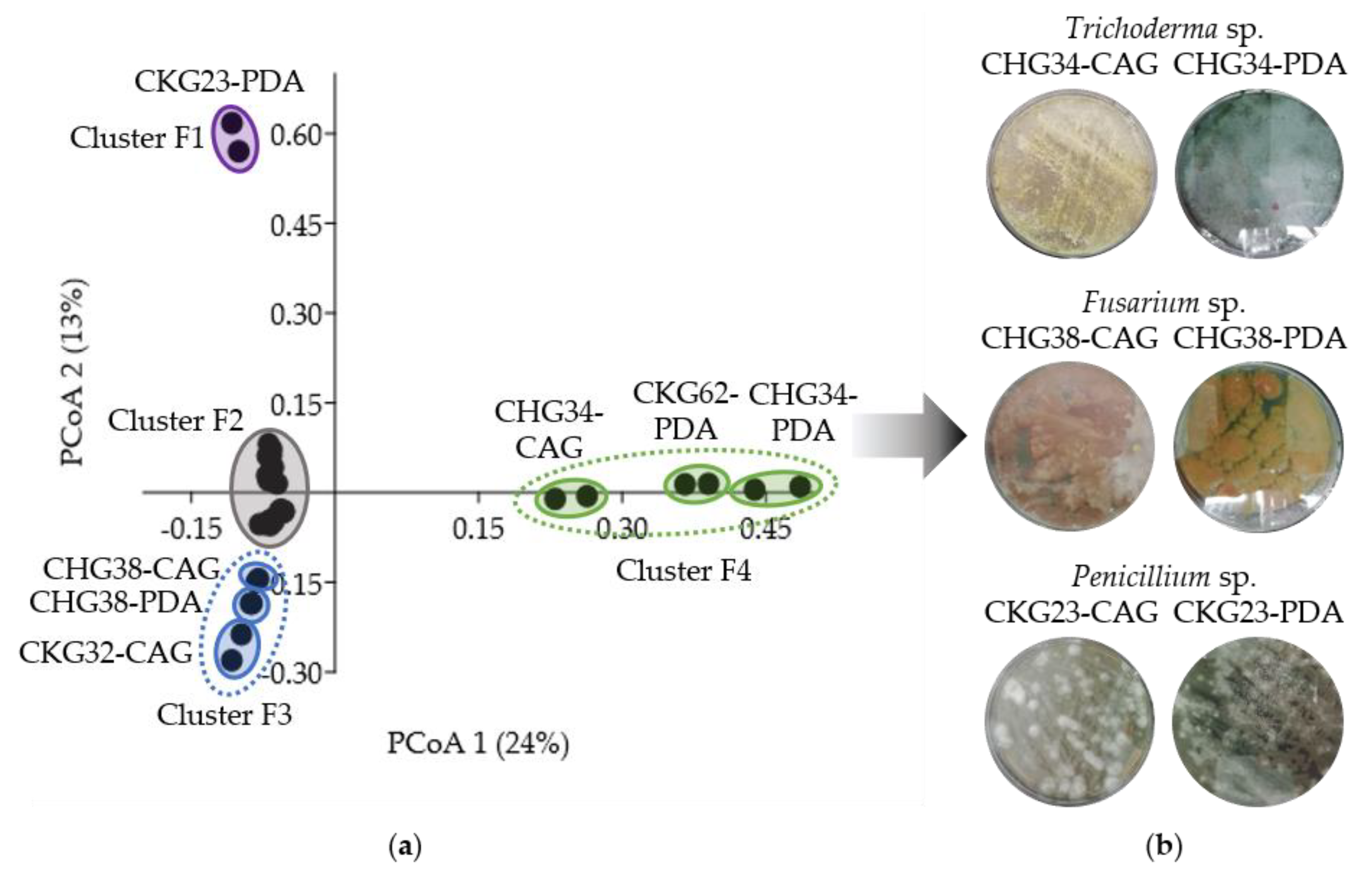
2.3. Bioactivity- and Metabolome-Based Selection of Microbial Extracts
2.4. IC50 Determinations of Prioritized Microbial Extracts
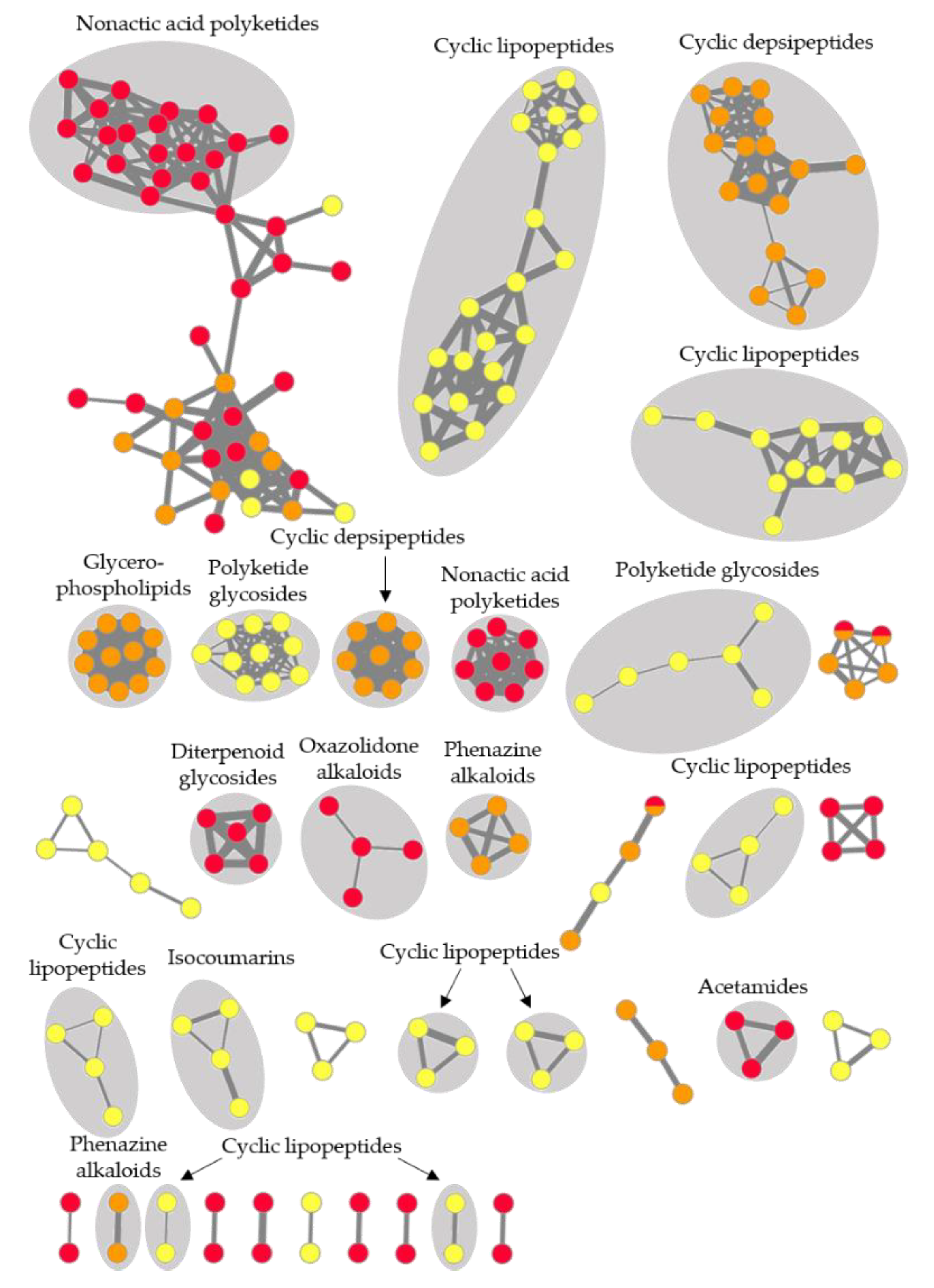
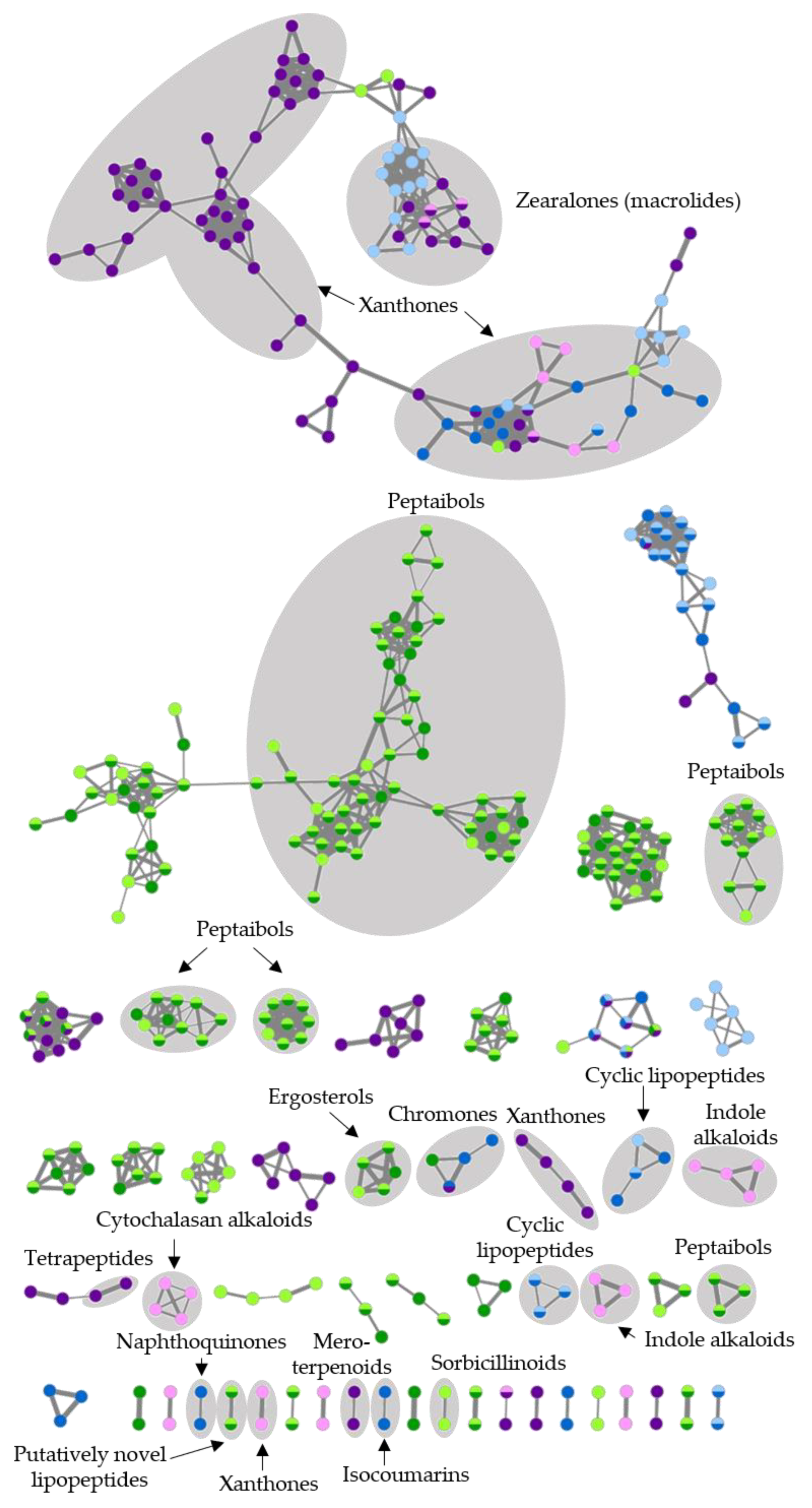
2.5. Feature-Based Molecular Networking and Dereplication of Nine Prioritized Microbial Extracts
3. Discussion
4. Materials and Methods
4.1. Sampling and Isolation of Microorganisms
4.2. DNA Extraction and Identification of Microbial Isolates
4.3. Cultivation and Extraction of Gut-Associated Microbial Strains
4.4. Biological Assays
4.5. Metabolomic Analyses
4.5.1. UPLC-QToF-MS/MS Measurements
4.5.2. Pre-Processing of UPLC-MS/MS Data and Statistics
4.5.3. Feature-Based Molecular Networking and Dereplication
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dishaw, L.J.; Cannon, J.P.; Litman, G.W.; Parker, W. Immune-directed support of rich microbial communities in the gut has ancient roots. Dev. Comp. Immunol. 2014, 47, 36–51. [Google Scholar] [CrossRef] [Green Version]
- Fraune, S.; Anton-Erxleben, F.; Augustin, R.; Franzenburg, S.; Knop, M.; Schröder, K.; Willoweit-Ohl, D.; Bosch, T.C.G. Bacteria–bacteria interactions within the microbiota of the ancestral metazoan Hydra contribute to fungal resistance. ISME J. 2014, 9, 1543–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Nunez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Westerdahl, A.; Olsson, J.C.; Kjelleberg, S.; Conway, P.L. Isolation and characterization of turbot (Scophtalmus maximus)-associated bacteria with inhibitory effects against Vibrio anguillarum. Appl. Environ. Microbiol. 1991, 57, 2223–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugita, H.; Matsuo, N.; Hirose, Y.; Iwato, M.; Deguchi, Y. Vibrio sp. strain NM 10, isolated from the intestine of a Japanese coastal fish, has an inhibitory effect against Pasteurella piscicida. Appl. Environ. Microbiol. 1997, 63, 4986–4989. [Google Scholar] [CrossRef] [Green Version]
- Sawabe, T.; Oda, Y.; Shiomi, Y.; Ezura, Y. Alginate degradation by bacteria isolated from the gut of sea urchins and abalones. Microb. Ecol. 1995, 30, 193–202. [Google Scholar] [CrossRef]
- Kobayashi, J.; Ishibashi, M. Bioactive metabolites of symbiotic marine microorganisms. Chem. Rev. 1993, 93, 1753–1769. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed marine natural products in the pharmaceutical and cosmeceutical industries: Tips for success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Hu, J.S.; Xu, J.L.; Shao, C.L.; Wang, G.Y. Biological and chemical diversity of ascidian-associated microorganisms. Mar. Drugs 2018, 16, 362. [Google Scholar] [CrossRef] [Green Version]
- Thomas, T.R.A.; Kavlekar, D.P.; LokaBharathi, P.A. Marine drugs from sponge-microbe association—A review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casertano, M.; Menna, M.; Imperatore, C. The ascidian-derived metabolites with antimicrobial properties. Antibiotics 2020, 9, 510. [Google Scholar] [CrossRef] [PubMed]
- Rath, C.M.; Janto, B.; Earl, J.; Ahmed, A.; Hu, F.Z.; Hiller, L.; Dahlgren, M.; Kreft, R.; Yu, F.; Wolff, J.J.; et al. Meta-omic characterization of the marine invertebrate microbial consortium that produces the chemotherapeutic natural product ET-743. ACS Chem. Biol. 2011, 6, 1244–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhao, S.; Ding, W.; Wang, P.; Yang, X.; Xu, J. Methylthio-aspochalasins from a marine-derived fungus Aspergillus sp. Mar. Drugs 2014, 12, 5124–5131. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhao, Z.; Ding, W.; Ye, B.; Wang, P.; Xu, J. Aspochalazine A, a novel polycyclic aspochalasin from the fungus Aspergillus sp. Z4. Tetrahedron Lett. 2017, 58, 2405–2408. [Google Scholar] [CrossRef]
- Xu, Y.; Huang, R.; Liu, H.; Yan, T.; Ding, W.; Jiang, Y.; Wang, P.; Zheng, D.; Xu, J. New polyketides from the marine-derived fungus Letendraea sp. 5XNZ4-2. Mar. Drugs 2019, 18, 18. [Google Scholar] [CrossRef] [Green Version]
- Alain, K.; Querellou, J. Cultivating the uncultured: Limits, advances and future challenges. Extremophiles 2009, 13, 583–594. [Google Scholar] [CrossRef]
- Garza, D.R.; Dutilh, B.E. From cultured to uncultured genome sequences: Metagenomics and modeling microbial ecosystems. Cell. Mol. Life Sci. 2015, 72, 4287–4308. [Google Scholar] [CrossRef] [Green Version]
- Tamaki, H.; Sekiguchi, Y.; Hanada, S.; Nakamura, K.; Nomura, N.; Matsumura, M.; Kamagata, Y. Comparative analysis of bacterial diversity in freshwater sediment of a shallow eutrophic lake by molecular and improved cultivation-based techniques. Appl. Environ. Microbiol. 2005, 71, 2162–2169. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Fu, C.; Wang, G. Microbial diversity associated with ascidians: A review of research methods and application. Symbiosis 2017, 71, 19–26. [Google Scholar] [CrossRef]
- Carver, C.E.; Mallet, A.L.; Vercaemer, B. Biological synopsis of the solitary tunicate Ciona intestinalis. Can. Man. Rep. Fish. Aquat. Sci. 2006, 2746, 1–55. [Google Scholar]
- Dishaw, L.J.; Flores-Torres, J.A.; Mueller, M.G.; Karrer, C.R.; Skapura, D.P.; Melillo, D.; Zucchetti, I.; De Santis, R.; Pinto, M.R.; Litman, G.W. A Basal chordate model for studies of gut microbial immune interactions. Front. Immunol. 2012, 3, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leigh, B.A.; Liberti, A.; Dishaw, L.J. Generation of germ-free Ciona intestinalis for studies of gut-microbe interactions. Front. Microbiol. 2016, 7, 2092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franchi, N.; Ballarin, L. Immunity in Protochordates: The tunicate perspective. Front. Immunol. 2017, 8, 674. [Google Scholar] [CrossRef]
- Dishaw, L.J.; Leigh, B.; Cannon, J.P.; Liberti, A.; Mueller, M.G.; Skapura, D.P.; Karrer, C.R.; Pinto, M.R.; De Santis, R.; Litman, G.W. Gut immunity in a protochordate involves a secreted immunoglobulin-type mediator binding host chitin and bacteria. Nat. Commun. 2016, 7, 10617. [Google Scholar] [CrossRef] [Green Version]
- Leigh, B.; Karrer, C.; Cannon, J.P.; Breitbart, M.; Dishaw, L.J. Isolation and characterization of a Shewanella phage-host system from the gut of the tunicate, Ciona intestinalis. Viruses 2017, 9, 60. [Google Scholar] [CrossRef]
- Liberti, A.; Cannon, J.P.; Litman, G.W.; Dishaw, L.J. A soluble immune effector binds both fungi and bacteria via separate functional domains. Front. Immunol. 2019, 10, 369. [Google Scholar] [CrossRef] [Green Version]
- Dishaw, L.J.; Flores-Torres, J.; Lax, S.; Gemayel, K.; Leigh, B.; Melillo, D.; Mueller, M.G.; Natale, L.; Zucchetti, I.; De Santis, R.; et al. The gut of geographically disparate Ciona intestinalis harbors a core microbiota. PLoS ONE 2014, 9, e93386. [Google Scholar] [CrossRef]
- Blasiak, L.C.; Zinder, S.H.; Buckley, D.H.; Hill, R.T. Bacterial diversity associated with the tunic of the model chordate Ciona intestinalis. ISME J. 2014, 8, 309–320. [Google Scholar] [CrossRef] [Green Version]
- Utermann, C.; Echelmeyer, V.A.; Blümel, M.; Tasdemir, D. Culture-dependent microbiome of the Ciona intestinalis tunic: Isolation, bioactivity profiling and untargeted metabolomics. Microorganisms 2020, 8, 1732. [Google Scholar] [CrossRef] [PubMed]
- Nothias, L.F.; Petras, D.; Schmid, R.; Duhrkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Allard, P.M.; Peresse, T.; Bisson, J.; Gindro, K.; Marcourt, L.; Pham, V.C.; Roussi, F.; Litaudon, M.; Wolfender, J.L. Integration of molecular networking and in-silico MS/MS fragmentation for natural products dereplication. Anal. Chem. 2016, 88, 3317–3323. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, L.; Montagu, M.; Landreau, A.; Mohamed-Benkada, M.; Grovel, O.; Sallenave-Namont, C.; Biard, J.F.; Amiard-Triquet, C.; Amiard, J.C.; Pouchus, Y.F. Peptaibols: Stable markers of fungal development in the marine environment. Chem. Biodivers. 2007, 4, 1116–1128. [Google Scholar] [CrossRef] [PubMed]
- Marik, T.; Tyagi, C.; Balazs, D.; Urban, P.; Szepesi, A.; Bakacsy, L.; Endre, G.; Rakk, D.; Szekeres, A.; Andersson, M.A.; et al. Structural diversity and bioactivities of peptaibol compounds from the Longibrachiatum Clade of the filamentous fungal fenus Trichoderma. Front. Microbiol. 2019, 10, 1434. [Google Scholar] [CrossRef] [Green Version]
- Paizs, B.; Suhai, S. Fragmentation pathways of protonated peptides. Mass Spectrom. Rev. 2005, 24, 508–548. [Google Scholar] [CrossRef]
- Da Silva, R.R.; Dorrestein, P.C.; Quinn, R.A. Illuminating the dark matter in metabolomics. Proc. Natl. Acad. Sci. USA 2015, 112, 12549–12550. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira Costa, O.; Nahas, E. Growth and enzymatic responses of phytopathogenic fungi to glucose in culture media and soil. Braz. J. Microbiol. 2012, 43, 332–340. [Google Scholar] [CrossRef] [Green Version]
- Goddard-Dwyer, M.; Lopez-Legentil, S.; Erwin, P.M. Microbiome variability across the native and invasive range of the ascidian Clavelina oblonga. Appl. Environ. Microbiol. 2020. AEM.02233-20. [Google Scholar] [CrossRef]
- Giatsis, C.; Sipkema, D.; Smidt, H.; Verreth, J.; Verdegem, M. The colonization dynamics of the gut microbiota in tilapia larvae. PLoS ONE 2014, 9, e103641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menna, M. Antitumor potential of natural products from Mediterranean ascidians. Phytochem. Rev. 2009, 8, 461–472. [Google Scholar] [CrossRef]
- Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics (World Health Organization 2017). Available online: https://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 19 October 2020).
- Chen, L.; Wang, X.-N.; Fu, C.-M.; Wang, G.-Y. Phylogenetic analysis and screening of antimicrobial and antiproliferative activities of culturable bacteria associated with the ascidian Styela clava from the Yellow Sea, China. Biomed. Res. Int. 2019, 2019, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanenko, L.A.; Kalinovskaya, N.I.; Mikhailov, V.V. Taxonomic composition and biological activity of microorganisms associated with a marine ascidian Halocynthia aurantium. Russ. J. Mar. Biol. 2001, 27, 291–296. [Google Scholar] [CrossRef]
- Meziti, A.; Kormas, K.A.; Pancucci-Papadopoulou, M.A.; Thessalou-Legaki, M. Bacterial phylotypes associated with the digestive tract of the sea urchin Paracentrotus lividus and the ascidian Microcosmus sp. Russ. J. Mar. Biol. 2007, 33, 84–91. [Google Scholar] [CrossRef]
- Laport, M.S.; Bauwens, M.; Collard, M.; George, I. Phylogeny and antagonistic activities of culturable bacteria associated with the gut microbiota of the sea urchin (Paracentrotus lividus). Curr. Microbiol. 2018, 75, 359–367. [Google Scholar] [CrossRef]
- Agrawal, S.; Acharya, D.; Adholeya, A.; Barrow, C.J.; Deshmukh, S.K. Nonribosomal peptides from marine microbes and their antimicrobial and anticancer potential. Front. Pharmacol. 2017, 8, 828. [Google Scholar] [CrossRef]
- Hifnawy, M.S.; Fouda, M.M.; Sayed, A.M.; Mohammed, R.; Hassan, H.M.; AbouZid, S.F.; Rateb, M.E.; Keller, A.; Adamek, M.; Ziemert, N.; et al. The genus Micromonospora as a model microorganism for bioactive natural product discovery. RSC Adv. 2020, 10, 20939–20959. [Google Scholar] [CrossRef]
- Macia-Vicente, J.G.; Shi, Y.N.; Cheikh-Ali, Z.; Grun, P.; Glynou, K.; Kia, S.H.; Piepenbring, M.; Bode, H.B. Metabolomics-based chemotaxonomy of root endophytic fungi for natural products discovery. Environ. Microbiol. 2018, 20, 1253–1270. [Google Scholar] [CrossRef]
- Oppong-Danquah, E.; Passaretti, C.; Chianese, O.; Blümel, M.; Tasdemir, D. Mining the metabolome and the agricultural and pharmaceutical potential of sea foam-derived fungi. Mar. Drugs 2020, 18, 128. [Google Scholar] [CrossRef] [Green Version]
- Crüsemann, M.; O’Neill, E.C.; Larson, C.B.; Melnik, A.V.; Floros, D.J.; da Silva, R.R.; Jensen, P.R.; Dorrestein, P.C.; Moore, B.S. Prioritizing natural product diversity in a collection of 146 bacterial strains based on growth and extraction protocols. J. Nat. Prod. 2016, 80, 588–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, H.B.; Bethe, B.; Hofs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. Chembiochem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Tison, D.L.; Kelly, M.T. Vibrio species of medical importance. Diagn. Microbiol. Infect. Dis. 1984, 2, 263–276. [Google Scholar] [CrossRef]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) principle to marine microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Lan, X.; Wang, Y.; Tian, L.; Fang, Y.; Zhang, L.; Zhang, K.; Zheng, X. New bioactive derivatives of nonactic acid from the marine Streptomyces griseus derived from the plant Salicornia sp. Phytochem. Lett. 2015, 12, 190–195. [Google Scholar] [CrossRef]
- Jeong, S.Y.; Shin, H.J.; Kim, T.S.; Lee, H.S.; Park, S.K.; Kim, H.M. Streptokordin, a new cytotoxic compound of the methylpyridine class from a marine-derived Streptomyces sp. KORDI-3238. J. Antibiot. 2006, 59, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Kusche, B.R.; Smith, A.E.; McGuirl, M.A.; Priestley, N.D. Alternating pattern of stereochemistry in the nonactin macrocycle is required for antibacterial activity and efficient ion binding. J. Am. Chem. Soc. 2009, 131, 17155–17165. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Ondeyka, J.; Guan, Z.; Dietrich, L.; Burgess, B.; Wang, J.; Singh, S.B. Isolation, structure and biological activities of platensimycin B4 from Streptomyces platensis. J. Antibiot. 2009, 62, 699–702. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Huo, P.; Chen, H.; Li, S.; Jiang, Y.; Li, L.; Xu, L.; Jiang, C.; Huang, X. New derivatives of nonactic and homononactic acids from Bacillus pumilus derived from Breynia fruticosa. Chem. Biodivers. 2014, 11, 1088–1098. [Google Scholar] [CrossRef]
- Oku, N.; Matoba, S.; Yamazaki, Y.M.; Shimasaki, R.; Miyanaga, S.; Igarashi, Y. Complete stereochemistry and preliminary structure-activity relationship of rakicidin A, a hypoxia-selective cytotoxin from Micromonospora sp. J. Nat. Prod. 2014, 77, 2561–2565. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Kunimoto, S.; Ikeda, D. Rakicidin A: A hypoxia-selective cytotoxin. Biol. Pharm. Bull. 2007, 30, 261–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landwehr, W.; Karwehl, S.; Schupp, P.J.; Schumann, P.; Wink, J. Biological active rakicidins A, B and E produced by the marine Micromonospora sp. isolate Guam1582. Adv. Biotechnol. Microbiol. 2016, 1, 1–5. [Google Scholar] [CrossRef]
- Charan, R.D.; Schlingmann, G.; Janso, J.; Bernan, V.; Feng, X.; Carter, G.T. Diazepinomicin, a new antimicrobial alkaloid from a marine Micromonospora sp. J. Nat. Prod. 2004, 67, 1431–1433. [Google Scholar] [CrossRef] [PubMed]
- Itoh, J.; Omoto, S.; Shomura, T.; Nishizawa, N.; Miyado, S.; Yuda, Y.; Shibata, U.; Inouye, S. Amicoumacin-A, a new antibiotic with strong antiinflammatory and antiulcer activity. J. Antibiot. 1981, 34, 611–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konda, Y.; Nakagawa, A.; Harigaya, Y.; Onda, M.; Masuma, R.; Omura, S. Aurantinin B, a new antimicrobial antibiotic from bacterial origin. J. Antibiot. 1988, 41, 268–270. [Google Scholar] [CrossRef] [Green Version]
- Zhuravleva, O.I.; Afiyatullov, S.S.; Ermakova, S.P.; Nedashkovskaya, O.I.; Dmitrenok, P.S.; Denisenko, V.A.; Kuznetsova, T.A. New C14-surfactin methyl ester from the marine bacterium Bacillus pumilus KMM 456. Russ. Chem. Bull. 2011, 59, 2137–2142. [Google Scholar] [CrossRef]
- Mhammedi, A.; Peypoux, F.; Besson, F.; Michel, G. Bacillomycin F, a new antibiotic of iturin group: Isolation and characterization. J. Antibiot. 1982, 35, 306–311. [Google Scholar] [CrossRef] [Green Version]
- Kimura, K.; Nakayama, S.; Nakamura, J.; Takada, T.; Yoshihama, M.; Esumi, Y.; Itoh, Y.; Uramoto, M. SNA-60-367, new peptide enzyme inhibitors against aromatase. J. Antibiot. 1997, 50, 529–531. [Google Scholar] [CrossRef] [Green Version]
- Mohamed-Benkada, M.; Montagu, M.; Biard, J.F.; Mondeguer, F.; Verite, P.; Dalgalarrondo, M.; Bissett, J.; Pouchus, Y.F. New short peptaibols from a marine Trichoderma strain. Rapid Commun. Mass Spectrom. 2006, 20, 1176–1180. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, P.K.; Wiest, A.; Ruiz, N.; Keightley, A.; Moran-Diez, M.E.; McCluskey, K.; Pouchus, Y.F.; Kenerley, C.M. Two classes of new peptaibols are synthesized by a single non-ribosomal peptide synthetase of Trichoderma virens. J. Biol. Chem. 2011, 286, 4544–4554. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.U.; Yun, B.S.; Lee, S.J.; Kim, J.H.; Yoo, I.D. Atroviridins A-C and neoatroviridins A-D, novel peptaibol antibiotics produced by Trichoderma atroviride F80317. I. Taxonomy, fermentation, isolation and biological activities. J. Antibiot. 2002, 55, 557–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawas, U.W.; Farrag, A.R.H.; Ahmed, E.F.; Abou El-Kassem, L.T. Cytotoxic effect of Fusarium equiseti fungus metabolites against N-Nitrosodiethylamine- and CCL4-induced hepatocarcinogenesis in rats. Pharm. Chem. J. 2018, 52, 326–333. [Google Scholar] [CrossRef]
- Wang, Q.X.; Bao, L.; Yang, X.L.; Guo, H.; Yang, R.N.; Ren, B.; Zhang, L.X.; Dai, H.Q.; Guo, L.D.; Liu, H.W. Polyketides with antimicrobial activity from the solid culture of an endolichenic fungus Ulocladium sp. Fitoterapia 2012, 83, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Holenstein, J.; DÉFago, G. Inheritance of naphthazarin production and pathogenicity to pea in Nectria haematococca. J. Exp. Bot. 1983, 34, 927–935. [Google Scholar] [CrossRef]
- Shiono, Y.; Tsuchinari, M.; Shimanuki, K.; Miyajima, T.; Murayama, T.; Koseki, T.; Laatsch, H.; Funakoshi, T.; Takanami, K.; Suzuki, K. Fusaristatins A and B, two new cyclic lipopeptides from an endophytic Fusarium sp. J. Antibiot. 2007, 60, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, A.E. Fusarium Mycotoxins: Chemistry, Genetics, and Biology; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2006. [Google Scholar]
- Utermark, J.; Karlovsky, P. Role of zearalenone lactonase in protection of Gliocladium roseum from fungitoxic effects of the mycotoxin zearalenone. Appl. Environ. Microbiol. 2007, 73, 637–642. [Google Scholar] [CrossRef] [Green Version]
- Pompeo, M.M.; Cheah, J.H.; Movassaghi, M. Total synthesis and anti-cancer activity of all known communesin alkaloids and related derivatives. J. Am. Chem. Soc. 2019, 141, 14411–14420. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Chen, H.; Li, W.; Zhu, X.; Ding, W.; Li, C. Bioactive chaetoglobosins from the mangrove endophytic fungus Penicillium chrysogenum. Mar. Drugs 2016, 14, 172. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, K.F.; Dalsgaard, P.W.; Smedsgaard, J.; Larsen, T.O. Andrastins A-D, Penicillium roqueforti metabolites consistently produced in blue-mold-ripened cheese. J. Agr. Food Chem. 2005, 53, 2908–2913. [Google Scholar] [CrossRef]
- Sigwart, J.D.; Blasiak, R.; Jaspars, M.; Jouffray, J.-B.; Tasdemir, D. Unlocking the potential of marine biodiscovery. Nat. Prod. Rep. 2021. [Google Scholar] [CrossRef]
- Davison, E.K.; Brimble, M.A. Natural product derived privileged scaffolds in drug discovery. Curr. Opin. Chem. Biol. 2019, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Oppong-Danquah, E.; Parrot, D.; Blümel, M.; Labes, A.; Tasdemir, D. Molecular networking-based metabolome and bioactivity analyses of marine-adapted fungi co-cultivated with phytopathogens. Front. Microbiol. 2018, 9, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Silber, J.; Ohlendorf, B.; Labes, A.; Erhard, A.; Imhoff, J.F. Calcarides A-E, antibacterial macrocyclic and linear polyesters from a Calcarisporium strain. Mar. Drugs 2013, 11, 3309–3323. [Google Scholar] [CrossRef] [PubMed]
- Utermann, C.; Parrot, D.; Breusing, C.; Stuckas, H.; Staufenberger, T.; Blümel, M.; Labes, A.; Tasdemir, D. Combined genotyping, microbial diversity and metabolite profiling studies on farmed Mytilus spp. from Kiel Fjord. Sci. Rep. 2018, 8, 7983. [Google Scholar] [CrossRef] [PubMed]
- Gomes, N.C.; Fagbola, O.; Costa, R.; Rumjanek, N.G.; Buchner, A.; Mendona-Hagler, L.; Smalla, K. Dynamics of fungal communities in bulk and maize rhizosphere soil in the tropics. Appl. Environ. Microbiol. 2003, 69, 3758–3766. [Google Scholar] [CrossRef] [Green Version]
- Hopple, J.S., Jr.; Vilgalys, R. Phylogenetic relationships in the mushroom genus Coprinus and dark-spored allies based on sequence data from the nuclear gene coding for the large ribosomal subunit RNA: Divergent domains, outgroups, and monophyly. Mol. Phylogenet. Evol. 1999, 13, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Wiese, J.; Labes, A.; Kramer, A.; Schmaljohann, R.; Imhoff, J.F. Lindgomycin, an unusual antibiotic polyketide from a marine fungus of the Lindgomycetaceae. Mar. Drugs 2015, 13, 4617–4632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, R.B. Mycology Guidebook; University of Washington Press: Seattle, WA, USA, 1974; p. 682. [Google Scholar]
- Pfeifer Barbosa, A.L.; Wenzel-Storjohann, A.; Barbosa, J.D.; Zidorn, C.; Peifer, C.; Tasdemir, D.; Cicek, S.S. Antimicrobial and cytotoxic effects of the Copaifera reticulata oleoresin and its main diterpene acids. J. Ethnopharmacol. 2019, 233, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.E.; Marner, M.; Labes, A.; Tasdemir, D. Rapid metabolome and bioactivity profiling of fungi associated with the leaf and rhizosphere of the Baltic seagrass Zostera marina. Mar. Drugs 2019, 17, 419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Oresic, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- van Santen, J.A.; Jacob, G.; Singh, A.L.; Aniebok, V.; Balunas, M.J.; Bunsko, D.; Neto, F.C.; Castaño-Espriu, L.; Chang, C.; Clark, T.N.; et al. The Natural Products Atlas: An open access knowledge base for microbial natural products discovery. ACS Cent. Sci. 2019, 5, 1824–1833. [Google Scholar] [CrossRef] [Green Version]
- Djoumbou-Feunang, Y.; Pon, A.; Karu, N.; Zheng, J.; Li, C.; Arndt, D.; Gautam, M.; Allen, F.; Wishart, D.S. CFM-ID 3.0: Significantly improved ESI-MS/MS prediction and compound identification. Metabolites 2019, 9, 72. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Identification | MRSA | Efm | Ab | Ec | Kp | Psa | Ca | Cn |
| CHG48-GYM | Streptomyces sp. | 5.0 | 4.5 | >100 | >100 | >100 | >100 | 12.9 | 13.1 |
| CKG20-GYM | Micromonospora sp. | 10.3 | 0.1 | >100 | >100 | >100 | >100 | >100 | >100 |
| CKG24-GYM | Bacillus sp. | 0.4 | 2.0 | >100 | >100 | >100 | >100 | >100 | 17.1 |
| CHG34-CAG | Trichoderma sp. | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 |
| CHG34-PDA | Trichoderma sp. | 36.8 | 32.0 | >100 | >100 | >100 | >100 | 3.7 | 58.8 |
| CHG38-CAG | Fusarium sp. | 4.0 | 3.0 | >100 | >100 | >100 | >100 | 9.9 | 20.9 |
| CHG38-PDA | Fusarium sp. | 10.8 | 7.6 | >100 | >100 | >100 | >100 | 11.4 | >100 |
| CKG23-CAG | Penicillium sp. | 19.8 | 29.8 | 4.9 | 15.6 | 8.9 | 15.8 | >100 | >100 |
| CKG23-PDA | Penicillium sp. | 8.3 | 18.1 | 42.5 | 41.0 | 31.4 | 42.6 | >100 | >100 |
| Positive control | 3.1 | 0.4 | 0.02 | 1.4 | 0.4 | 0.4 | 1.3 | 0.1 |
| Extract | Identification | A375 | A549 | HCT116 | MB231 |
| CHG48-GYM | Streptomyces sp. | 5.8 | 0.02 | 21.4 | 22.1 |
| CKG20-GYM | Micromonospora sp. | 0.8 | 1.6 | 1.3 | 1.4 |
| CKG24-GYM | Bacillus sp. | 70.7 | 67.9 | 86.6 | >100 |
| CHG34-CAG | Trichoderma sp. | 67.9 | 70.1 | 71.9 | 69.7 |
| CHG34-PDA | Trichoderma sp. | 19.3 | 31.1 | 24.1 | 32.8 |
| CHG38-CAG | Fusarium sp. | 35.9 | 70.1 | 62.1 | >100 |
| CHG38-PDA | Fusarium sp. | 92.3 | >100 | >100 | >100 |
| CKG23-CAG | Penicillium sp. | 2.0 | 4.9 | 8.5 | 2.5 |
| CKG23-PDA | Penicillium sp. | 5.2 | 17.5 | 27.6 | 9.0 |
| Positive control | 0.8 | 1.3 | 13.3 | 2.3 |
| Extract | Identification | Nodes | Putatively Annotated Chemical Families | Annotation Rate (%) |
| CHG48-GYM | Streptomyces sp. | 73 | Acetamides, diterpenoid glycosides, linear polyketides, nonactic acid polyketides, oxazolidone alkaloids | 24 |
| CKG20-GYM | Micromonospora sp. | 61 | Cyclic depsipeptides, phenazine alkaloids, glycerophospholipids | 40 |
| CKG24-GYM | Bacillus sp. | 89 | Cyclic lipopeptides, isocoumarins, polyketide glycosides | 71 |
| CHG34-CAG | Trichoderma sp. | 173 | Ergosterols, peptaibols, sorbicillinoids | 27 |
| CHG34-PDA | Trichoderma sp. | 171 | ||
| CHG38-CAG | Fusarium sp. | 53 | Chromones, cyclic lipopeptides, isocoumarins, naphthoquinones, xanthones, zearalenones | 38 |
| CHG38-PDA | Fusarium sp. | 54 | ||
| CKG23-CAG | Penicillium sp. | 29 | Cytochalasans, indole alkaloids, mero- and sesquiterpenoids, styrylpyrones, tetrapeptides, xanthones, zearalenones | 48 |
| CKG23-PDA | Penicillium sp. | 108 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Utermann, C.; Echelmeyer, V.A.; Oppong-Danquah, E.; Blümel, M.; Tasdemir, D. Diversity, Bioactivity Profiling and Untargeted Metabolomics of the Cultivable Gut Microbiota of Ciona intestinalis. Mar. Drugs 2021, 19, 6. https://doi.org/10.3390/md19010006
Utermann C, Echelmeyer VA, Oppong-Danquah E, Blümel M, Tasdemir D. Diversity, Bioactivity Profiling and Untargeted Metabolomics of the Cultivable Gut Microbiota of Ciona intestinalis. Marine Drugs. 2021; 19(1):6. https://doi.org/10.3390/md19010006
Chicago/Turabian StyleUtermann, Caroline, Vivien A. Echelmeyer, Ernest Oppong-Danquah, Martina Blümel, and Deniz Tasdemir. 2021. "Diversity, Bioactivity Profiling and Untargeted Metabolomics of the Cultivable Gut Microbiota of Ciona intestinalis" Marine Drugs 19, no. 1: 6. https://doi.org/10.3390/md19010006