
Abstract
The southern African subcontinent and its surrounding oceans accommodate globally unique ecoregions, characterized by exceptional biodiversity and endemism. This diversity is shaped by extended and steep physical gradients or environmental discontinuities found in both ocean and terrestrial biomes. The region’s biodiversity has historically been the basis of life for indigenous cultures and continues to support countless economic activities, many of them unsustainable, ranging from natural resource exploitation, an extensive fisheries industry and various forms of land use to nature-based tourism.
Being at the continent’s southern tip, terrestrial species have limited opportunities for adaptive range shifts under climate change, while warming is occurring at an unprecedented rate. Marine climate change effects are complex, as warming may strengthen thermal stratification, while shifts in regional wind regimes influence ocean currents and the intensity of nutrient-enriching upwelling.
The flora and fauna of marine and terrestrial southern African biomes are of vital importance for global biodiversity conservation and carbon sequestration. They thus deserve special attention in further research on the impacts of anthropogenic pressures including climate change. Excellent preconditions exist in the form of long-term data sets of high quality to support scientific advice for future sustainable management of these vulnerable biomes.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- Southern Africa
- Marine biomes
- Terrestrial biomes
- Ecoregions
- Endemism
- Biogeography
- Upwelling
- Climate change
- Science–policy interface
- Biodiversity conservation
- Cross-realm
- Anthropogenic impacts
- Interdisciplinary research
1 Introduction
Southern Africa is a globally important hotspot of biodiversity and endemism. It hosts both individual taxa and entire ecological communities that are unique in the world. Its large marine biomes, while also containing many unique, endemic species, are known more for their exceptionally high biomass and productivity than for their overall species richness. For the purposes of this chapter, we have defined southern Africa as the area south of the Kunene River on the West Coast and Quelimane on the East Coast (approximately 17° S). As such, it incorporates all of Namibia, South Africa, Botswana, Eswatini (formerly Swaziland) and Lesotho, as well as most of Zimbabwe and the southern half of Mozambique. Aspects of the marine Angola Current biome between the Congo River (approximately 6°S) and the Kunene River are also included.
Identified as one of the world’s 17 megadiverse nations, South Africa ranks in the top ten nations globally for plant species richness and is third for marine species endemism (Tolley et al. 2019). This is despite the fact that it is located mostly outside of tropical latitudes which host most of the world’s species-rich ecosystems. With a landmass of 1.2 million km2 and surrounding seas of 1.1 million km2, South Africa (without its sub-Antarctic territories and waters) is among the smaller of the world’s megadiverse countries—which together contain more than two-thirds of the world’s biodiversity (Tolley et al. 2019). Tolley et al. (2019) reported that approximately 10% of the world’s marine fish species, 7% of vascular plants, 5% of mammals, 7% of birds, 4% of reptiles, 2% of amphibians and 1% of freshwater fishes exist in South Africa. While they found limited information available on invertebrate groups overall, they stated that almost a quarter of global cephalopod species (octopus, squid and cuttlefish) are found in South African waters.
Southern African marine ecosystems offer habitats for a large variety of species. Even though species richness is lower than in the top-ranked ecosystems such as coral reefs, they host a high proportion of globally unique taxa. Griffiths et al. (2010) reported 12,715 marine eukaryotes in the ocean around South Africa, and the National Biodiversity Assessment in 2018 lists more than 13,000 marine species (Sink et al. 2019). However, these estimates constantly need updating owing to new species discoveries and taxonomic revisions. Many of the marine species in South African waters have so far been found nowhere else on the globe, and thus, the country is reported as having the third highest marine endemism rate after New Zealand (51%) and Antarctica (45%) (Sink et al. 2019). Accurate estimates of biodiversity and endemism are challenging to obtain, and records differ because studies vary in areal extent covered, methodology, data sources consulted, sampling effort and taxa investigated. Griffiths and Robinson (2016) concluded, based on then available comprehensive data sets for marine species, that around 28% to 33% of the marine taxa within the national boundaries of South Africa are endemic. It was, however, pointed out that sampling intensity varies greatly among habitats, and has been concentrated in the shallow near-shore areas. South African benthic invertebrates show peaks in species richness (regardless of a likely bias in sampling intensity), where species distribution ranges overlap when two biogeographic regions meet, particularly around the Cape Peninsula, where the influences of the Indian Ocean and the Atlantic Ocean converge (Awad et al. 2002). The region between Cape Point and Cape Agulhas, a transition zone between the cool-temperate West Coast and the warm-temperate South Coast biogeographic provinces, hosts distinct genetic lineages of several species which are unique to this zone (Teske et al. 2011). Maximum marine endemism has been found in the warm temperate Agulhas ecoregion along the South Coast of South Africa (Awad et al. 2002; Sink et al. 2019). However, several species like the African penguin, green sea turtle, abalone and many fish taxa are currently endangered due to habitat losses and other drivers.
The terrestrial ecosystems of southern Africa have a very rich biodiversity, particularly considering the subcontinent’s low rainfall and subtropical-to-temperate climate. South Africa holds three of the world’s 35 biodiversity hotspots (a measure of biological diversity combined with vulnerability to threats): the Cape Floristic region, Succulent Karoo Biome and Maputaland–Pondoland–Albany center of endemism (Tolley et al. 2019). There are an estimated 22,000 species of ferns, angiosperms and gymnosperms in southern Africa (Huntley 2003). Species diversity is not homogeneous and varies extensively throughout the region. In the southern African terrestrial habitats, floral speciation and endemism are particularly high within the Fynbos and Succulent Karoo Biomes. The succulent karoo vegetation represents the greatest floral species richness when compared globally to all areas with equivalent rainfall. Although the savanna has only slightly fewer plant species than the Fynbos Biome, the nature of the diversity is very different. Firstly, the southern African savanna is an order of magnitude greater in spatial extent than the fynbos. Secondly, there is a high alpha diversity, but low beta and gamma diversity in the savanna. This means there is a high diversity of species at any particular location, but these same species occur very widely (Huntley 2003). The fynbos, by contrast, has relatively few species at any specific location, that is a lower alpha diversity, but high differences in species between locations, thus high beta and gamma diversity (Cowling et al. 2003a; Scholes 2003). Plant speciation at the genus level is truly exceptional within the fynbos, with genera such as Protea and Erica showing immense and relatively recent speciation (Cowling et al. 2003b) along with Aizoaceae in the succulent karoo (Klak et al. 2004).
Faunal diversity is in part explained by the diverse flora (Procheş and Cowling 2006). Terrestrial animal diversity, although less than plant diversity, is still high. About 960 species of birds, of which 98 are endemic, occur in the region (van Rensburg et al. 2002). Southern Africa has the richest reptile diversity in Africa, exceeding 490 species (Branch 2006) with 384 indigenous species in South Africa, Lesotho and Eswatini (Branch 2014), and including a diversity hotspot region for chameleons (Tolley et al. 2008). Insect diversity, though still not fully studied, is closely correlated with plant species diversity (Procheş and Cowling 2006). Spider diversity is higher than in the African tropics and 71 spider families, 471 genera and 2170 species have been recorded from South Africa, of which 60% are endemic (Dippenaar-Schoeman et al. 2015).
Spatially extended and sometimes steep environmental gradients are thought to be largely responsible for the extensive biome-level diversity on land (Cowling et al. 2003b). These authors point out that high species richness is often related to dystrophic soils and disturbance, which prevents the establishment of shaded closed woodland and forest conditions even under high rainfall, and appears to promote high plant richness and endemism.
Deeper in time, the relative stability of major climatic and ocean features over the Quaternary and late Neogene periods has likely lowered extinction rates relative to many other parts of the world (Enquist et al. 2019). Over the Quaternary period, repeated glacial/interglacial cycles have caused pulsed equatorward/poleward shifts in the path of westerly rain-bearing frontal systems to the subcontinent and shifts in the position of trade winds (Stuut et al. 2004) and in sea level, exposing coastal shallows and platforms for periods of thousands to tens of thousands of years (Cowling et al. 2020).
Unlike on land, a certain degree of environmental stability appears to be a key factor promoting high biodiversity in the marine realm (Woodd-Walker et al. 2002; Robison 2004, 2009). Highly fluctuating coastal upwelling conditions on the southern African West Coast thus reduce biodiversity. In contrast, the interplay of the warm-water Agulhas Current with the cold-water Benguela Current at the southern coast sections as well as more stable conditions at the East Coast create high numbers of heterogeneous micro- and macrohabitats that enhance biodiversity and endemism.
Sharp spatial discontinuities shape the marine realm, namely via the topography, major ocean currents and wind-driven circulation affecting water properties—particularly temperature, salinity, oxygen and nutrient content—which determine the habitat structure and living conditions of organisms. One such major environmental discontinuity is generated by the convergence of the warm and saline Agulhas Current, which originates from the Indian Ocean and moves poleward (southward) along the East Coast of southern Africa, the South Atlantic Current and the cold eutrophic branches of the Antarctic Circumpolar Current from the south, feeding into the Benguela Current which flows equatorward (northward) along the West Coast of southern Africa (see Figs. 8.1 and 8.2; Chap. 8 and Fig. 9.5; Chap. 9).
Southern Africa may well be the area in which modern humans evolved (Fortes-Lima et al. 2022). The history of human impacts in this region is extensive. Small numbers of modern humans persisted in the southern Cape, in particular, during very adverse climatic conditions of Pleistocene stadials, supplementing their terrestrial diets with marine resources (Esteban et al. 2020; Wren et al. 2020), but only with likely local impacts. Coastal marine resources such as mussels and limpets were hand-harvested along the South African South Coast for food and other uses during the Middle and Late Stone Ages (Nelson-Viljoen and Kyriacou 2017). The San, a predominantly hunter-gatherer society, and the Khoekhoen, early pastoralists, have inhabited the region going back 150,000 to 260,000 years (Schlebusch et al. 2017). Bantu-speaking groups migrated into southern Africa by both a West African and an East African route, with settlements dating back approximately 1400 years (Vansina 1994; van Waarden 2002). These groups brought with them crop agriculture, metal smelting and cattle. Finally, there was European colonization of the area starting from the fifteenth century, including Portuguese settlements on the East Coast and Dutch settlements in the Cape region (Biggs and Scholes 2002). During the Holocene, human populations increased in size, and population growth of indigenous peoples and European colonization characterize the past millennium to century time scales. Tidal fish traps were in use from the late nineteenth century (Hine et al. 2010), before the advent of industrial-scale fishing for sole and Cape hakes in the early twentieth century (Durholtz et al. 2015). In the past few centuries, the introduction of modern technologies into the region has had sudden, massive impacts on both land and ocean biodiversity (Skowno et al. 2019). In the terrestrial environment, European settlers introduced guns to the area, which had devastating impacts on large mammals, especially elephants. In addition, many areas were destocked of their original wildlife, which was replaced with domestic livestock, often in fenced areas with artificial water points. Later there was further extensive transformation of natural vegetation to agricultural cropland.
Southern Africa has extensive and well-maintained legacy environmental data. For instance, the South African National Biodiversity Institute (SANBI) has the responsibility for coordinating the maintenance of national biodiversity information (www.sanbi.org/resources/infobases/). The South African Earth Observation Network (SAEON) maintains long-term environmental monitoring sites in South Africa (Chap. 30). Although at a slightly smaller scale, comprehensive research and data collections regarding biodiversity are also maintained in Namibia, Botswana, Zimbabwe and Mozambique. Much of the southern African historic data is well maintained and accessible in electronic format. Avian biodiversity is exceptionally well monitored through annual citizen science bird censuses (Hugo and Altwegg 2017).
2 Oceanic Biomes
The biogeography of the oceans is structured primarily through large-scale ocean circulation patterns, where frontal zones between water masses act as boundaries (Longhurst 2007). Biomes and provinces within the oceans have been defined using different classification methods (Hardman-Mountford et al. 2008; Oliver and Irwin 2008). Considering the oceans’ biogeochemical properties, Fay and McKinley (2014) have distinguished 17 open-ocean biomes. Other authors based their biogeographic zonation of the oceans on a combination of physical and biogeochemical properties, as well as ecological communities (compare bioregions defined in Lombard et al. 2004; Sutton et al. 2017). Briggs and Bowen (2012) based their classification on fish distributions and defined the warm-temperate Benguela Province as a separate zone within the East Atlantic, and also the Agulhas Province, which is linked to the Indo-Pacific. In this chapter, we adopt a broad definition of ‘oceanic biomes’ for the marine realm, including not only abiotic criteria, but also ecological zonation patterns.
2.1 Oceanographic Gradients Shaping Southern African Marine Biomes
Satellite images and satellite-derived data products visualize some of the key characteristics of the marine biomes around southern Africa (Fig. 2.1; Good et al. 2020). On the West Coast, at about 17°S, the Angola-Benguela Frontal Zone (ABFZ) separates the warm oligotrophic waters off Angola from the Benguela Upwelling System (BUS) to the south, which is characterized by cold and nutrient-rich surface waters. In contrast, all along the East Coast of Africa, warm waters are found at the surface. The warm-water masses of the Angola Current in the north, and of the Agulhas Current in the south-east, form the oceanographic boundaries of the cold BUS (Hutchings et al. 2009; Kirkman et al. 2016; Chap. 9).

Sea surface temperature (SST) around southern Africa; mean for the period 2003–2019. AC Angola Current, ABFZ Angola-Benguela Frontal Zone, BCC Benguela Coastal Current, PUC Poleward Undercurrent, BOC Benguela Offshore Current, SEJ Shelf Edge Jet, ACR Agulhas Current Retroflection, ARC Agulhas Return Current. Data: OSTIA product (https://resources.marine.copernicus.eu/product-detail/SST_GLO_SST_L4_REP_OBSERVATIONS_010_011/INFORMATION), DOI: https://doi.org/10.48670/moi-00168
The circulation in Angolan waters is dominated by the Angola Current (AC) which transports warm tropical waters southward until reaching the ABFZ (Kopte et al. 2018). South of the ABFZ, the subsurface Poleward Undercurrent (PUC) transports South Atlantic Central Water (SACW) poleward, providing warmer pelagic habitats in the otherwise cold Benguela environment. South-easterly trade winds of the South Atlantic Ocean drive the northward-flowing Benguela Current and the coastal upwelling, with surface waters being colder than the surrounding water masses. At the surface, the Benguela Current is a key dynamic feature, and branches into an oceanic part (Benguela Offshore Current, BOC) and a coastal part (Benguela Coastal Current, BCC). The BOC is part of the eastern limb of the Subtropical Gyre transporting subtropical Eastern South Atlantic Central Waters (ESACW) northward. The BCC can be seen as a coastal jet being part of the upwelling system (Siegfried et al. 2019).
The currents east and south of southern Africa provide an important part of the large-scale circulation in the Indian Ocean and establish a key link of the global conveyor belt circulation. As the western boundary current in this system, the Agulhas Current transports water of tropical and subtropical origin to the south. It flows along the South African South Coast, then overshoots the southern tip of the continent, before abruptly turning back into the Indian Ocean. At the Agulhas Current retroflection, large mesoscale eddies are shed which, together with small-scale currents like the Shelf Edge Jet, make the Cape Cauldron south-east of Africa a region of strong current interactions and vigorous air–sea exchange. As a result, the Agulhas waters provide a warm and saline component for the upper limb of the Atlantic Meridional Overturning Circulation (AMOC), and are important for global climate (Beal et al. 2011). Some of these waters also reach into the Benguela region (Fig. 2.1; for details on the Agulhas Current System, see Chap. 8).
2.2 Southern African Marine Biomes: A Brief Overview
Around southern Africa, different marine biomes or bioregions (sensu Lombard et al. 2004) can be distinguished, which are defined through their oceanographic properties, as well as by their biological communities. Five inshore bioregions have been described for the coast of South Africa alone (Lombard et al. 2004), namely (from east to west) the Delagoa, Natal, Agulhas, South-western Cape and the Namaqua bioregions (Fig. 2.2), the latter bordering the Namib bioregion off Namibia (Griffiths et al. 2010). As offshore bioregions, Lombard et al. (2004) defined the Atlantic Offshore, the Indo-Pacific, the West Indian Offshore and the South-west Indian Offshore Bioregions (see Fig. 2.2). In their spatial biodiversity assessment for South Africa, those authors further divided their five inshore and four offshore bioregions by depth strata into ‘biozones,’ units which may help to assess threat status and design protection measures.

South Africa’s five inshore and four offshore bioregions, as defined by Lombard et al. (2004); from Griffiths et al. (2010), https://doi.org/10.1371/journal.pone.0012008.g004
Within the Benguela region or province, a description of ecologically meaningful spatial subunits has been summarized by Hutchings et al. (2009) and Kirkman et al. (2016). Recently, researchers have advised taking these boundaries into account to implement ecosystem-based management and effective conservation measures (Kirkman et al. 2016, 2019). The concept of marine biomes is also being integrated into current efforts to implement Marine Spatial Planning (MSP) in the southern African countries (see Sect. 2.2.4 in this chapter).
The marine ecosystems around the southern African coasts cover the transition zone between Atlantic and Indio-Pacific biomes (Fig. 2.3). This gives rise to a remarkably rich biodiversity and a high amount of endemic species. For South Africa alone, more than 3500 species were classified as endemic during the Census of Marine Life, within a total of 12,715 reported eukaryotic species (Costello et al. 2010). The marine realm around the coasts of Angola, Namibia, South Africa and Mozambique comprises a large variety of habitats, including sandy and rocky shores, kelp forests, coral reefs and estuaries. Yet, an uneven sampling effort, in both geographical and taxonomic dimensions, as well as differential availability of region-specific identification guides and expertise, are expected to cause bias in the reported biodiversity (Griffiths 2005; Costello et al. 2010).

Marine biomes and major current systems around southern Africa. The warm Angola and Agulhas Currents and the cold Benguela Current shape the ecosystems around the coasts of southern Africa. The four major biogeographic provinces (cool temperate, warm temperate, subtropical and tropical) each host characteristic assemblages of species. According to Teske et al. (2011), ‘coastal phylogeographic breaks’ (between Cape Point and Cape Agulhas, between Algoa Bay and the Wild Coast, and around St Lucia) characterize zones where distinct genetic lineages can be found in many coastal taxa. LME—Large Marine Ecosystem; sBUS and nBUS—southern and northern Benguela subsystem, respectively. (Figure adapted from Teske et al. 2011)
Depending on their distance from shore and their depth, the marine ecosystems can generally be grouped into three broad zones, namely coastal, benthic and pelagic ecosystems. For the West Coast of southern Africa, these zones have already been used in ecosystem mapping for South Africa’s National Biodiversity Assessment (Sink et al. 2011), and in extending the approach across the national borders to cover the entire Benguela region (Kirkman et al. 2019). Alongshore, the southern African marine ecosystems are associated with the major biomes of the Angola Current region and the Benguela Upwelling System (BUS)—which together form the Benguela Current Large Marine Ecosystem (BCLME). The BCLME extends from the northern boundary of Angola southward along the West Coast of southern Africa to the Cape of Good Hope in South Africa. The BUS hosts a cell of exceptionally intense upwelling off Lüderitz (around 27°S), which effectively divides the BUS into a northern and a southern part (Duncombe Rae 2005; Shannon 2009). Together with the adjacent Agulhas Current LME, these zones belong to the biogeochemical provinces BENG (Benguela Current coast) and EAFR (East African coast), respectively, according to Longhurst (2007).
Aside from their horizontal zonation with distance from shore, the southern African marine biomes are vertically structured and distinct: near-coastal biomes are clearly influenced by the contact with the seafloor and its benthic communities, and furthermore to varying degrees by river run-off or transport of water and minerals between land and sea (see Sect. 2.4 of this chapter). Further offshore, the steep slope of the continental shelf forms an important structure—shaping upwelling intensity, ocean currents and thereby also migration and dispersal routes for organisms. Far offshore, the open ocean is mainly structured by the diminishing intensity of light with increasing depth, leading to the typical oceanic zones: (1) the photic epipelagic zone reaching from the surface down to approximately 200 m (the zone with the largest productivity, based on photosynthesis), (2) the mesopelagic twilight zone from ~200 to 1000 m (inhabited also by vertically migrating species, which seek refuge here from predators during the day and ascend into the epipelagic at night to feed), (3) the bathypelagic zone (1000 to 4000 m) with no sunlight and no primary producers at all, and (4) the abyssopelagic zone (below 4000 m) with highest ambient pressure and temperatures close to 0°C. The ecology of the latter two zones of the global ocean is least studied, mainly due to the technical and logistical requirements to reach them with oceanographic instrumentation.
2.2.1 Angola Current Biome
The ecosystem properties north of the Angola-Benguela Frontal Zone (ABFZ), which is located at about 17°S, are driven by coastal trapped waves propagating southward from the Equatorial Atlantic, thereby resulting in seasons of dynamically driven upwelling and downwelling. Water is supplied from the open South Atlantic toward the eastern boundary by the South Equatorial Undercurrent and the South Equatorial Counter Current and transported southward along the coast by the Angola Current. Additionally, freshwater is supplied by river run-off from the Congo River and other rivers further south, such as the Cuanza River (Kirkman et al. 2016, and references therein, Siegfried et al. 2019). There is no significant wind-driven upwelling north of the ABFZ. Variable extents of low-oxygen waters occur and create an oxygen-depleted subsurface zone encompassing the Angola Dome region and extending toward the coast (Monteiro and van der Plas 2006). The main exploited resources of this subsystem include two Sardinella species and Kunene horse mackerel Trachurus trecae as pelagic fish, as well as demersal sparid fish such as Dentex spp., Angolan hake Merluccius polli, and deep-sea red crab Chaceon sp. (Kirkman et al. 2016). The biodiversity of demersal species in Angola is high, relative to the temperate Benguela ecosystem to the south (Huntley et al. 2019; Kirkman and Nsingi 2019).
In the steep environmental gradient at the ABFZ, high abundances of zooplankton (Verheye et al. 2001; Postel et al. 2007) and fish larvae (Ekau et al. 2001) have been observed. This front constitutes the northern boundary of the BCLME, which is divided into two oceanographically and ecologically distinct regions, the northern and the southern Benguela Upwelling subsystems.
2.2.2 The Northern Benguela Upwelling System (nBUS)
The northern Benguela Upwelling System (nBUS) extends from the Angola-Benguela Frontal Zone in the north (17oS) to the Lüderitz upwelling cell off Namibia in the south (27oS). Upwelling in the nBUS is perennial, with total cumulative upwelling being an order of magnitude greater than in the sBUS (Lamont et al. 2018). Extreme low-oxygen or even anoxic zones develop on the inner continental shelf off Namibia, due to a combination of factors: rather oxygen-poor SACW reaching the nBUS, and high oxygen consumption coupled with high sulfate reduction rates at the sea floor (microbial respiration and fermentation of the decaying organic matter deposited onto the shelf sediments). Hydrogen sulfide (H2S) is maintained in large (1500 km2) bacterial mats produced by sulfur-oxidizing, nitrate- and sulfur-storing bacteria, which prevent H2S from constantly diffusing into the water column. However, gas (methane and H2S) eruptions from this layer lead to seasonally occurring intrusions of hydrogen sulfide, with detrimental effects, including mass mortalities, on the surrounding marine organisms (Emeis et al. 2004; Brüchert et al. 2006; Currie et al. 2018).
Stock sizes of formerly important fisheries resources in the nBUS, specifically sardine (Sardinops sagax) and, to a lesser extent, anchovy (Engraulis encrasicolus), collapsed in the 1970s. Nowadays, abundant components of the food web include large scyphozoan jellyfishes (Aequorea forskalea and Chrysaora fulgida) (Roux et al. 2013), the bearded goby (Sufflogobius bibarbatus) (Utne-Palm et al. 2010) and mesopelagic fishes. Top predators include Cape fur seals (Arctocephalus pusillus pusillus), dolphins and seabirds, e.g., African penguins (Spheniscus demersus) and Cape gannets (Morus capensis) (Kirkman et al. 2016). Commercially exploited components include predatory demersal species—specifically the two hake species (Merluccius paradoxus and M. capensis) and monkfish (Lophius vomerinus)—and pelagic horse mackerel (Trachurus capensis) as well as deep-sea red crab (Chaceon maritae) (Kirkman et al. 2016).
2.2.3 The Southern Benguela Upwelling System (sBUS)
South of the permanent Lüderitz upwelling cell, in the southern Benguela Upwelling System (sBUS), low-oxygen water is less prevalent than in the nBUS and occurs in rather restricted locations. The sBUS is characterized by seasonal, wind-driven upwelling at discrete centers, with peaks occurring during austral spring and summer (Hutchings et al. 2009; Lamont et al. 2018). The densest communities of zooplankton have been reported downstream of the Namaqua and Cape Columbine upwelling cells, between the Orange River mouth and St Helena Bay (Pillar 1986; Huggett et al. 2009), in the major nursery grounds of commercially targeted fish species. The southern section of the sBUS is influenced by the Agulhas Current and Agulhas leakage into the South Atlantic, which occurs through shedding of mesoscale Agulhas rings and eddies (Beal et al. 2011). The landside boundary between the Benguela Current LME and the Agulhas Current LME is located at Cape Agulhas. The seaward transition, on the other hand, moves within certain limits as the variable mesoscale oceanographic features of the Agulhas leakage lead to different degrees of influence and exchange around the boundary between the two Large Marine Ecosystems.
The continental shelf of the Benguela region is particularly wide compared to the other eastern boundary upwelling systems. Thus, in conjunction with shelf-break fronts originating from the Shelf Edge Jet and inshore upwelling fronts, retention cells are formed, particularly in the sBUS, increasing the productivity (Flynn et al. 2020; Rixen et al. 2021). They facilitate nurseries for anchovies and other small pelagic fish species spawned west of Cape Agulhas as well as for mesopelagic pseudo-oceanic species, and provide rich feeding grounds for whales (Ragoasha et al. 2019; Dey et al. 2021).
Exploited marine resources in the sBUS include hakes, anchovy, sardine, a currently declining overexploited West Coast rock lobster (Jasus lalandii) population, linefish such as snoek (Thyrsites atun) and yellowtail (Seriola lalandii), and several tuna species (Kirkman et al. 2016). There is considerable exchange between the stocks of small pelagics of the sBUS and the Agulhas Current LME. Commercial and experimental fishing on mesopelagic resources has occurred in the sBUS where catches were dominated by the lanternfish Lampanyctodes hectoris (Tyler 2016), while this species as well as lightfishes (Maurolicus walvisensis, formerly reported as M. muelleri) were found to be common and abundant over the shelf of the entire BUS (Coetzee et al. 2009, 2018).
2.2.4 Agulhas Current LME
The Agulhas Current Large Marine Ecosystem stretches from Cape Agulhas to the northern end of the Mozambique Channel, and the ecosystem is driven by the swift southward-moving warm Agulhas Current. Intermittent upwelling occurs at the shelf edge and seasonal mixing takes place on the broad Agulhas Bank (Kirkman et al. 2016, and references therein). Strong winds from various directions are typical for the area. The Agulhas Bank provides spawning grounds for many commercially important species targeted by fisheries locally or after their migration to the West Coast. These include the hakes, sardine, anchovy, round herring (Etrumeus whiteheadi), horse mackerel, chokka-squid (Loligo reynaudii), Agulhas sole (Austroglossus pectoralis) and linefish, such as dusky kob (Argyrosomus japonicus) (Kirkman et al. 2016). Seabirds, particularly Cape gannets, are top predators of the region’s pelagic fishes. The zooplankton community on the Agulhas Bank is dominated by a large copepod species, Calanus agulhensis (Verheye et al. 1994; Huggett et al. 2023), which is an important food item for pelagic fishes and squid. Highest copepod densities are often associated with a quasi-permanent ridge of cool upwelled water on the central and eastern parts of the bank that is thought to fuel local productivity as well as enhance retention (Huggett and Richardson 2000). Around the South Coast of South Africa in the Agulhas Current LME, biodiversity is particularly high due to the influence of the Indian Ocean biota (Gibbons and Hutchings 1996; Smit et al. 2017).
2.3 The Benguela Upwelling System: A Focus Region of SPACES Research
Research within the TRAFFIC Footnote 1 project of the SPACES II Footnote 2 program has set a focus on the Benguela Upwelling System (BUS), in order to increase our understanding of biodiversity, ecological functioning, carbon sequestration and particularly trophic transfer efficiency within its food web, and hence the mechanisms supporting the exceptionally high productivity of this ocean region.
2.3.1 A Global Perspective on the Ecological Significance of the Benguela Region
Together with the Humboldt, Canary and California Current Systems, the Benguela Current System belongs to the world’s most productive marine biomes and provides a significant portion of the global catch of wild fishes for human consumption. Common to the biomes in all four of these large Eastern Boundary Upwelling Systems (EBUS) is a rich supply of nutrients through coastal upwelling caused by the trade winds, which supports intense phytoplankton growth that forms the base for an exceptionally high marine productivity. At the same time, the Benguela Upwelling System (BUS) hosts communities with a high share of globally unique (i.e., endemic) marine taxa.
The Benguela Current Large Marine Ecosystem (BCLME) has been included in the Tentative List of UNESCO World Heritage Sites and has been classified as one of Outstanding Universal Value (OUV) based on three criteria: (1) the extremely high primary production sustaining a global hotspot of productivity and rich stocks of commercially targeted fish and crustaceans, as well as large populations of other fishes, seabirds and marine mammals; (2) the occurrence of many endemic or otherwise rare species; and (3) the massive genesis of seabird guano, used as natural fertilizer to enhance agriculture production.
2.3.2 Biome-Level Diversity
The communities within the marine biomes around southern Africa are shaped by both spatial boundaries and seasonal processes, which generate dynamic environmental gradients within the ocean.
The high productivity of the marine ecosystem in the Benguela region results from the process of wind-driven coastal upwelling, which defines the specific oceanographic conditions on the shelf and over the continental rise. Upwelling occurs seasonally in short (~10 day) cycles in the sBUS, while it is perennial in the nBUS (Hutchings et al. 2009; Lamont et al. 2018). During austral summer, hypoxic, nutrient-rich South Atlantic Central Water (SACW) from the Angola Gyre is transported into the northern Benguela, whereas during the winter season the oxygen-rich Eastern SACW (ESACW) spreads northward (Monteiro and van der Plas 2006; Monteiro et al. 2006; Mohrholz et al. 2008).
Because the Benguela Current Upwelling System has been a focus region of research in SPACES, its key elements are highlighted in the following. For more details see Chap. 11. Figures 2.4 and 2.5 below highlight selected taxa observed in the BUS during SPACES research cruises. Further taxa are depicted in Chap. 11.

Selected species of mesopelagic invertebrates caught during SPACES cruise SO285 with the Research Vessel Sonne in 2021 (cruise report: https://doi.org/10.48433/cr_so285). Top left: cephalopod Histioteuthis bonnellii, top right: amphipod Themisto gaudichaudii, bottom left: a decapod shrimp, bottom right: a euphausiid (krill). Images not to same scale. © Solvin Zankl www.solvinzankl.com

Selected species of mesopelagic fishes caught during SPACES cruise SO285 with the Research Vessel Sonne in 2021 (cruise report: https://doi.org/10.48433/cr_so285). Top left: Maurolicus walvisensis, top right: Diaphus hudsoni, middle left: M. walvisensis—photophores as seen from below; middle right: Diaphus dumerilii; bottom left: meso- and bathypelagic angler fish Melanocetus johnsonii, bottom right: Stomias boa. Images not to same scale. © Solvin Zankl www.solvinzankl.com
2.3.2.1 Primary Production and Lower Trophic Levels
Upwelling brings nutrient-rich water from below the thermocline to the surface, providing nutrients for the growth of planktonic algae. As demonstrated for other regions (Ayón et al. 2008), plankton abundance and production increase under moderate upwelling conditions, both seasonally and locally (Grote et al. 2007; Bode et al. 2014). Strong turbulence during intense upwelling can hinder the primary production as phytoplankton cells are swirled out of the euphotic surface water or advected offshore. In addition, abundance and productivity are initially low in freshly upwelled water, because this water originates below the thermocline and its phytoplankton content is minimal. Therefore, the development of a diatom-dominated phytoplankton bloom in the nutrient-rich upwelling plume takes time to respond to upwelling conditions.
Microzooplankton, organisms within the size range from 20 to 200 μm, are distributed in a clear shelf-to-offshore zonation. Heterotrophic dinoflagellates prevail in cold, recently upwelled waters on the shelf, whereas in warmer waters at the shelf break, small copepod species dominate the microzooplankton. For protists, naked ciliates, small dinoflagellates and tintinnids, a clear preference has been shown for the warmer water masses surrounding an upwelling filament in the nBUS. In contrast, the copepods Microsetella spp., Oithona spp. and Oncaea spp. were associated with the cold water inside the filament (Bohata 2015).
2.3.2.2 Mesozooplankton
Zooplankton play a vital role in the functioning of marine ecosystems, providing the main energy pathway from primary producers to higher trophic levels, contributing significantly to carbon sequestration via the biological pump, and serving as sentinels of climate change (Richardson 2008; Batten et al. 2019). Organisms between 200 μm and 2 cm in size make up the mesozooplankton. As in other marine ecosystems, copepods dominate the mesozooplankton of the Benguela Upwelling System in terms of abundance, biomass and diversity. Consequently, they have been the most intensely studied zooplankton group compared to other taxa (Shannon and Pillar 1986; Verheye et al. 1992; Bode et al. 2014).
Zooplankton diversity generally decreases from the equator toward the poles and increases from the epipelagic toward the meso- and bathypelagic zones which reach from 200 to 1000 and from 1000 m to 4000 m depth, respectively (Woodd-Walker et al. 2002; Kosobokova et al. 2011; Bode et al. 2018). Since diversity in the ocean is usually connected to moderate levels of ecosystem disturbance (Woodd-Walker 2001; Woodd-Walker et al. 2002), zooplankton diversity in upwelling zones tends to be relatively low, increasing with distance from shore. This has also been shown for the BUS (Gibbons and Hutchings 1996). Typical copepod species of the Benguela upwelling community include the medium to large Centropages brachiatus, Calanoides natalis (previously C. carinatus), Metridia lucens and Nannocalanus minor, as well as several smaller members of the Paracalanidae and Clausocalanidae families (De Decker 1964, 1984). De Decker (1984) observed that copepod diversity in the nBUS and sBUS increased from less than 20 species over the shelf to more than 20 or 30 species typically found farther offshore in the South Atlantic central gyre. A patch of up to 47 species off the sBUS were likely transported within Agulhas rings originating from the warm Agulhas Current. An increase in zooplankton diversity from west to east around the coast of South Africa, i.e., toward the Indian Ocean, was also noted by Gibbons and Hutchings (1996), as species with temperate affinities give way to subtropical communities.
A major research task of the TRAFFIC project was to compare the two physically and biologically contrasting subsystems, the nBUS and sBUS, in order to better understand how zooplankton dynamics, community structure and functional traits influence food-web structures at higher trophic levels, and to disentangle biological from physicochemical effects. Predominantly herbivorous species such as the common copepod C. natalis become very abundant and play a key role in the food web during active upwelling events. The population size of this species is regulated through a complex mechanism involving offshore displacement of older life-cycle stages in maturing upwelled water plumes and temporary developmental arrest (diapause) of pre-adults at greater depth during prolonged periods of starvation. The latter coincides with the non-upwelling period and the diapausing individuals return shoreward into the productive surface zones with the onset of the next upwelling season (Verheye et al. 1991; Verheye et al. 2005). C. natalis clearly dominates the upwelling regions of the nBUS and the sBUS, while Calanus agulhensis, the dominant large copepod on the Agulhas Bank off the South African coast (De Decker et al. 1991; Verheye et al. 1994; Huggett et al. 2023), occurs at lower abundances in the sBUS (Huggett and Richardson 2000), and only more recently and sporadically in the nBUS (Rittinghaus 2021). Apart from spatial differences, there are also marked seasonal variations in the zooplankton of the BUS, which are closely coupled to the seasonality of the upwelling cycle. Besides the limited number of studies on non-copepod taxa, the distribution, dynamics and taxonomic composition of smaller calanoid and cyclopoid copepod species, which fit well into the prey-size spectra for larvae or juveniles of many fish species, are understudied (Verheye et al. 2016).
Since the 1950s to 1960s, substantial long-term changes occurred in the abundance, biomass and production of individual zooplankton species. Simultaneously, species and size composition of neritic (on the shelf) mesozooplankton communities have shifted in both the nBUS and sBUS subsystems (Huggett et al. 2009; Bode et al. 2014; Verheye et al. 2016). Abundances of neritic copepods have increased during recent decades by at least one order of magnitude in both subsystems, with turning points reached around the mid-1990s in the south and around the mid-2000s in the north, after which they declined. At the same time, there were marked changes in the copepod community structure, with a gradual shift in dominance from larger to smaller species in both subsystems. These major long-term changes in zooplankton communities are likely to have fundamental effects on biogeochemical processes, food-web structure and ecosystem functioning of the BUS as well as on the ecosystem services, such as fisheries, that ultimately rely on the zooplankton (Verheye et al. 2016). Researchers are currently investigating how this change in community size structure is related to climate change on the one hand, and to changes in the predation regime on the other, due to fluctuating planktivorous pelagic fish populations (e.g., sardine and anchovy).
2.3.2.3 Macrozooplankton and Jellyfishes
Macrozooplankton, organisms between 2 and 20 cm body length, provide an important link between higher and lower trophic levels and serve as the primary food for many species, particularly fish but also seabirds and marine mammals. They also play an important role in the export of carbon from the surface to the intermediate and deep ocean (Moriarty et al. 2013). In the BUS, the diversity of pelagic decapods is high, with 46 of 91 Atlantic species present (Schukat et al. 2013; Sutton et al. 2017). Euphausiids, especially Euphausia hanseni, represent a major portion of the macrozooplankton in the Benguela system, and their biomass peaks near the shelf edge; also large swarms of Nyctiphanes capensis may assemble above the shelf (Barange and Stuart 1991; Hutchings et al. 1991; Werner and Buchholz 2013).
Jellyfishes (e.g., Chrysaora fulgida and Aequorea forskalea) can be very abundant, particularly in the northern Benguela (Roux et al. 2013). Yet, jellyfishes and comb jellies (ctenophores) have been understudied (Brodeur et al. 2016; Gibbons et al. 2021), because of their poor quantitative representation in plankton nets. However, their role in the food web should receive further attention, especially since they have repeatedly increased in abundance under the influence of climate change and adverse fishing regimes (Lynam et al. 2006; Roux et al. 2013; Brodeur et al. 2016; Opdal et al. 2019). In the nBUS, jellyfishes appear to lead to dead-end food chains, since they have very limited nutritional value to top predators. An acceleration of this so-called jellification process has been hypothesized, should pronounced oxygen-minimum zones expand further under climate change (Ekau et al. 2018).
2.3.2.4 Cephalopods
Another characteristic of the pelagic food web is a high diversity of cephalopods (squids and cuttlefishes, specifically) which also distinguish the fauna of the Benguela region (and the Agulhas Current with its retroflection) from other regions of the Atlantic (Rosa et al. 2008). According to Tolley et al. (2019), South Africa hosts almost 25% of the world’s cephalopod species. The ecoregions along the temperate coasts of southern Africa are inhabited by numerous species of cuttlefish, bobtails and squids, while they feature a rather low diversity of octopuses (Rosa et al. 2019). The role of cephalopods in the food webs around southern Africa has not yet been thoroughly assessed and requires further research, as already noted by Shannon et al. (2003). Therefore, within the TRAFFIC project, a combination of methods—microscopy, biochemical and genetic analyses—have been applied in order to investigate cephalopod feeding behaviour and their predatory interactions within the food webs of the Benguela upwelling subsystems.
2.3.2.5 Pelagic Fish Species
The Benguela ecosystem hosts a species spectrum typical of ‘wasp-waist’ upwelling systems, where a low number of so-called ‘small pelagics’ (the ‘wasp-waist’ with sardine, anchovy and round herring) control both the lower trophic levels (zooplankton) and the higher trophic levels such as tuna or hake and also seabirds and marine mammals (Cury et al. 2000). The small pelagic fishes are characterized by strong fluctuations in their stock sizes and the capacity to build up large stock sizes within a few years (Schwartzlose et al. 1999). Both the northern and the southern Benguela underwent multiple regime shifts since the 1950s caused by both environmental and anthropogenic factors leading to a significant reduction in demersal and pelagic catches in recent years (Jarre et al. 2013, 2015; Heymans and Tomczak 2016). In the northern Benguela, a particularly intense fishery for sardines collapsed around 50 years ago and stocks have not recovered since (Kainge et al. 2020). Sardine and particularly anchovy are still caught in the southern Benguela subsystem, but these forage fish species in the northern subsystem have been practically replaced by horse mackerel, bearded goby and shallow-water hake in the far north and by bearded goby, Hector’s lanternfish (Lampanyctodes hectoris) and shallow-water hake in the southern part of the northern Benguela (Mwaala 2022).
2.3.2.6 Mesopelagic Fish Species
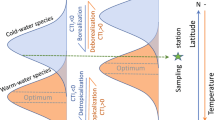
Mesopelagic fishes form an ecologically important component of the pelagic ecosystem, due to their biomass, their diversity, and their diel migrations, which greatly affect the vertical transport of carbon in the ocean (see Fig. 2.5). In the southern part of the sBUS, the proximity of the frontal system to the shelf break and prevailing retention cells, provide sufficient habitat for pseudo-oceanic species associated with continental shelf regions (Hulley and Lutjeharms 1989; Sutton et al. 2017). According to the latter authors, the landward extension of the distribution range of oceanic species and the seaward range extension of pseudo-oceanic species in the central and southern BUS (28–35°S) largely coincide with the 800-m isobath.
Hector’s lanternfish Lampanyctodes hectoris (Myctophidae) and lightfish Maurolicus walvisensis (Sternoptychidae) have previously been shown to be the most abundant mesopelagic fishes in the region (Hulley and Prosch 1987). Because knowledge about the ecology and diversity of mesopelagic fishes was still limited, this group received particular attention in the TRAFFIC project. Spatially, communities differed between the nBUS and sBUS, as well as between on-the-shelf and offshore within each subsystem. These assemblages contained both tropical warm-water species and cold-water species as well as pseudo-oceanic species such as L. hectoris, which was prominent on the shelf of the sBUS (Duncan et al. 2022). During a research cruise in late austral summer (Feb-Mar 2019), those authors reported 88 mesopelagic fish species of 22 families in the two subsystems of the Benguela Upwelling System. The most diverse families were lanternfishes (Myctophidae) with 35 species, followed by Stomiidae (ten species) and Sternoptychidae (eight species). About half of all specimens caught were Diaphus hudsoni (Myctophidae), M. walvisensis (Sternoptychidae) and Lampanyctus australis (Myctophidae) (Duncan et al. 2022; Chaps. 3 and 11).
2.3.2.7 Demersal Fish Species
The most important nursery grounds for pelagic spawners, as well as for a wide variety of demersal and predatory fishes, are located on the shelf areas of northern-central Namibia, the West Coast of South Africa, the Agulhas Bank and in the small but significant KwaZulu-Natal Bight on the East Coast of South Africa (Hutchings et al. 2002, 2009). Eggs and larvae from spawning grounds on the western Agulhas Bank are transported alongshore to the West Coast in the strong Shelf Edge Jet, keeping them close to the shallower shelf regions, rather than dispersing them offshore (Grote et al. 2012).
Two species of hake co-occur in the Benguela, Merluccius capensis and M. paradoxus. The former is typically found in shallower zones, the latter at greater depths, leading to their common names of shallow-water and deep-water hake, respectively. As important fisheries resources, these species have been well studied. In general, hakes occupy high trophic levels in the food web. The diet of the shallow-water hake changes throughout their lifetime toward an almost exclusive fish prey and includes common cannibalistic feeding behavior as well as substantial predation on its con-generic M. paradoxus. In contrast, the diet of the deep-water hake consists of about one half of crustaceans and one half of fish prey, even for large adult specimens. The deep-water hake therefore usually appears at a lower trophic level than the shallow-water hake in the northern Benguela (Wilhelm et al. 2015), and also in the southern Benguela (van der Lingen and Miller 2014; Durholtz et al. 2015). Other commercially important demersal fish resources found in the Benguela are monkfish (Lophius vomerinus), which are especially important in the northern Benguela, kingklip (Genypterus capensis) and Agulhas sole (Austroglossus pectoralis) and West Coast sole (A. microlepis).
2.3.2.8 Functioning of Marine Food Webs in the Benguela Upwelling System
A classical food chain that develops from upwelling events is comprised of large-sized diatoms, followed by large-sized zooplankton (herbivorous euphausiids and copepods), which are consumed by small pelagic fishes. Long-term studies of the plankton communities off the West Coast of southern Africa revealed an increase in mesozooplankton abundance, accompanied by a shift to smaller-sized plankton taxa, especially copepods, since the 1950s–1960s and until the mid-1990s in the sBUS and the mid-2000s in the nBUS, respectively (Verheye et al. 2016). Those authors concluded that the observed changes in copepod abundance and size structure could be attributed to the complex interplay of local warming or cooling, increased primary production where upwelling intensified, combined with reduced predation pressure by pelagic fishes owing to increased fishing. Verheye et al. (2016) also emphasized that there is uncertainty about the relative importance of these bottom-up and top-down forcing mechanisms. Overall, the classic picture of a direct coupling between upwelling intensity and primary and secondary production with a short food chain cannot be sustained in the nBUS (Ekau et al. 2018). Secondary (and primary) production is highest either temporarily during moderate upwelling conditions (Cushing 1996) or spatially at some distance from the upwelling source. This confirms the ‘optimal environmental window’ hypothesis of Cury and Roy (1989), stating that larval fish survival and fish recruitment are dependent on upwelling intensity in a dome-shaped function in Ekman-type upwellings, where very low and very high wind speeds are detrimental. Furthermore, longer trophic pathways than previously thought are active in the food web of the nBUS, contributing to a higher complexity and thus lower transfer efficiency of biomass and energy from phytoplankton to fish (Schukat et al. 2014). This may partly explain the differences in fish production between individual Eastern Boundary Upwelling Systems, and specifically between the nBUS and sBUS despite similar primary production rates (Chavez and Messié 2009).
2.3.3 Productivity and Resource Utilization
Being an Eastern Boundary Upwelling System, its core characteristics provide the Benguela ecosystem with an exceptionally high productivity across the food chain from plankton to top predators, which lays the foundation for an effective provision of living marine resources and the development of large fisheries.
The Benguela region is inhabited by a variety of taxa, which are typical of upwelling systems and some of the fish species can attain high biomass levels. Several species are commercially important as fisheries resources including Cape hakes (Merluccius capensis and M. paradoxus), Cape and Cunene horse mackerels (Trachurus capensis and T. trecae, respectively), and small pelagics (sardine Sardinops sagax, anchovy Engraulis encrasicolus and round herring Etrumeus whiteheadi). Despite their smaller catches Crustaceans such as rock lobster (Jasus lalandii) and deep-water red crab (Chaceon sp.) are also commercially important (van der Lingen et al. 2006c; Kirkman et al. 2016; Kainge et al. 2020; Chap. 11).
In terms of their economic value, the most important fisheries in the nBUS subsystem are those for Cape hakes (most valuable) and horse mackerel (largest volume), whereas in the sBUS, Cape hakes (most valuable) and small pelagic species (largest volume) dominate the fisheries (Kainge et al. 2020; Chap. 11). Additionally, high biomasses of as yet not commercially targeted mesopelagic fishes occur in the Benguela region, and gobies as well as jellyfishes can also appear at high abundances, particularly in the nBUS (Lynam et al. 2006; Roux et al. 2013; Kirkman et al. 2016; Salvanes et al. 2018). The sBUS—from south of Lüderitz to Cape Agulhas—provides nursery grounds for most of that subsystem’s ecologically and economically important fish species (Kirkman et al. 2016), including both hake species and small pelagics (Clupeiformes).
By the middle of the last century (1950 to late-1960s), sardine was the central fisheries resource in both subsystems, with peak catches beyond 1 million tons in the nBUS and close to half a million tons in the sBUS (for details see Chap. 11). Yet sardine catches declined rapidly due to overfishing, both off the Namibian and the South African coasts (van der Lingen et al. 2006c; Augustyn et al. 2018). In the sBUS, anchovy replaced sardine during the following three decades. In the nBUS, the collapse of the sardine population resulted in substantial reductions in the number of purse-seiners and labour-intensive canning factories and fish reduction plants, resulting in job losses for several thousands of people (Boyer and Oelofsen 2004), and the Namibian sardine fishery has essentially been replaced by the fishery for Cape horse mackerel.
The collapse of the sardine population has resulted in substantial ecosystem changes arising from changed trophic interactions, since sardine was the dominant forage fish species of a wide variety of predators, including other fishes, marine mammals and seabirds. In the nBUS, that change has been hypothesized to have promoted the proliferation of jellyfishes (Roux et al. 2013), and predators previously reliant on sardine as the main very fatty and nutritious food source, now utilize alternative food sources including bearded goby, lanternfish, horse mackerel, shallow-water hake and jellyfish. Goby appears to be a successful substitute when the other more nutritious species (horse mackerel in the north and lanternfish in the south) are not available, likely because of its high abundance in the system and tolerance to high temperature and low oxygen levels (Erasmus et al. 2021). In contrast, catches of the major South African fisheries have remained more or less stable over the past 70 years and pelagic fish are still abundant in the system. However, the most recent decline in sardine is concerning from both fisheries and ecosystem perspectives, particularly for endangered seabirds such as the African penguin, a species whose breeding success (Crawford et al. 2008) and mortality (Robinson et al. 2015) are correlated with the abundance and distribution of sardine.
An experimental fishery for mesopelagic fishes using midwater trawling was initiated as part of the South African small pelagic fishery in 2011, when a catch of 7000 tons was taken. That experiment was resumed in 2018 and 2019 with around 5000 tons taken each year, but the relatively high cost of fishing coupled with a general downturn in both the anchovy and sardine fisheries has resulted in its cessation (DEFF 2020).
2.3.4 Organizational Efforts Geared to Protect Marine Biodiversity
In 2007, the three coastal nations Angola, Namibia and South Africa founded the Benguela Current Commission (BCC), which in 2013 became a permanent intergovernmental institution through the Benguela Current Convention. Its purpose is to protect marine biodiversity and to promote sustainable use of the natural resources in the Benguela region through an ecosystem approach to ocean governance. All three nations defined national and common ‘Ecologically and Biologically Significant Areas’ (EBSAs), which have been and will be submitted to the Convention on Biological Diversity (CBD) and which form the basis for the designation of new Marine Protected Areas (Harris et al. 2022).
The International Union for Conservation of Nature, IUCN, has defined ‘A Global Standard for the Identification of Key Biodiversity Areas’ (IUCN 2016). Key Biodiversity Areas (KBAs) are ‘Sites contributing significantly to the global persistence of biodiversity’, in terrestrial, freshwater and marine ecosystems. Such KBAs are proposed and identified from the bottom up by local experts or private and government organizations through National Coordination Groups (NCGs). In order to promote an understanding of the spatial distribution and risk status of biodiversity, O’Hara et al. (2019) have linked the distribution ranges and conservation status (based on IUCN protected area categories) for thousands of marine species in order to present global maps of extinction risk of marine biodiversity.
The South African NCG is hosted by the South African National Biodiversity Institute (SANBI). South Africa hosts an exceptionally high number of endemic terrestrial and marine species (global rank 3 on the national level; SANBI). The National Biodiversity Assessment published by SANBI monitors and reports on the state of biological diversity in South Africa, in order to support political strategies for the conservation of biodiversity (Skowno et al. 2019). In Namibia, the National Biodiversity Assessments are hosted and published by the Ministry of Environment and Tourism (MET), Multi-lateral Environmental Agreements Division, supported by the National Biodiversity Strategies and Action Plan 2 (NBSAP2) steering committee (e.g., MET 2018).
2.4 Marine Spatial Planning in Southern Africa
Ongoing Marine Spatial Planning (MSP) in the Benguela Current Large Marine Ecosystem (BCLME) involves Angola, Namibia and South Africa, since part of the LME lies in each of their respective Exclusive Economic Zones (EEZs). Through the Benguela Current Convention, all three nations promote the vision of sustaining human and ecosystem well-being in the BCLME. With their MSP initiative, the countries in the Benguela region are among the first African coastal states to implement MSP, particularly since they pursue a transboundary perspective in their planning. The regional MSP strategy was adopted by the Benguela Current Convention (BCC 2018). The project MARISMA is a partnership between the BCC, its member states Angola, Namibia and South Africa and the government of Germany in pursuit of the sustainable development of the Benguela Current Large Marine Ecosystem. The project’s goal is ‘to maximize socio-economic benefits while ensuring the safeguarding of the marine ecosystem’s health and maintenance of marine services provision’ (www.benguelacc.org/marisma), leading to the very recent launch of the BCC GeoData Portal (https://geodata.benguelacc.org/). All three nations have divided their waters into several distinct planning zones, based on human uses and ecological boundaries (Finke et al. 2020; Fig. 2.6). Each nation developed a baseline report describing the actual status of their waters. The BCC countries prioritize—to varying degrees—the following objectives (see Finke et al. 2020 for details):
-
Protection of biodiversity features of national, regional or global significance (focus on EBSAs);
-
Providing access to fishing grounds, while protecting key fish habitats from adverse effects by human use, including fisheries;
-
Securing mariculture locations;
-
Enabling exploration and promoting sustainable use of geological resources;
-
Guaranteeing maritime transport and disposal of dredge material;
-
Allocating space for military training activities;
-
Enabling responsible marine and coastal tourism;
-
Protecting underwater infrastructure, e.g., cables; and
-
Protecting maritime and underwater cultural heritage.

The marine spatial planning (MSP) areas of Angola, Namibia and South Africa. Black dashed outline around the Benguela Current LME (from Finke et al. 2020)
Finke et al. (2020) conclude that fostering ecosystem-based MSP in the context of strong economic growth agendas requires balanced and integrated governance and technical planning structures and processes. While the regional and national approaches taken are considered useful, their implementation will still need to overcome obstacles regarding funding, data needs, research data management, legislation and institutionalization (Finke et al. 2020).
3 Terrestrial Biomes
3.1 Environmental Gradients Shaping Terrestrial Biomes
Terrestrial biomes in southern Africa can be described simply in terms of the relative dominance by tree, shrub, grass and annual plant life forms (see Rutherford and Westfall 1986). The distribution and structure of these biomes are likely shaped by a mix of linked and independent gradients of climate, disturbance and substrate (geology and soils). Their distinct contemporary structure has evolved particularly during the Neogene, a period of declining atmospheric CO2, increasing aridity, cooling and intensifying climate seasonality. During the Holocene, major climatic gradients include, broadly, a general tropical to temperature thermal regime from north (warm/humid) to south (cool/dry), an aridity gradient from west (dry) to east (wet), and a seasonality gradient from south-west (winter rainfall) to north east (summer rainfall). These climates provide the thermal and moisture regimes that underpin vegetation structural and related disturbance regimes that determine biome distributions, with some putative interactive role of substrate (soil texture and nutrient status).
The existence of flammable vegetation even under relatively high rainfall conditions appears to override assumed climatic controls of the dominance of trees, and thus the distribution of forests and woodlands. For this reason, flammable shrubland, grassland and savanna biomes may be found under rainfall conditions that have the potential to support taller and more closed vegetation with higher leaf area index over vast regions of the subcontinent (Bond et al. 2005). The dystrophic sandy soils of the south-western coastal plains and Cape Fold Mountains, and the deep sands of the Kalahari respectively support flammable shrublands of the Fynbos Biome, and the extensive well drained arid and mesic savannas of the central Kalahari. Heavier clay-rich soils are associated with arid and semiarid karoo shrublands that stretch from the arid western regions of Namibia and northern South Africa to the more mesic Thicket and Forest biomes of the eastern seaboard.
Paleoclimatic shifts associated with glacial periods have tended to increase the influence of cold and wet westerly rain-bearing frontal systems, and permitted a northern expansion of the associated winter-rainfall shrublands, while lower atmospheric CO2 and aridity appears to have reduced the dominance of trees in savanna-dominated central southern Africa, and increased grass dominance.
3.2 Southern African Terrestrial Biomes
Southern Africa has a disproportionately high level of terrestrial diversity at the biome level. This is largely as a consequence of the climatic conditions and the intensity or frequency of disturbance. Within southern Africa nine uniquely different biome level vegetation structures have been identified (Figs. 2.7 and 2.8) (Olson et al. 2001; Rutherford et al. 2006). These are Forest, Fynbos, Succulent Karroo, Nama Karoo, Grassland, Savanna (including embedded halophytic pans and flooded grasslands), Thicket, Indian Ocean Tropical Belt (IOTB) and Desert. The Desert biome is the smallest biome in South Africa, but is better developed and extends northward along the Namibian coast to Angola. Despite this high biome diversity, it is savanna woodlands which dominate in southern African countries. These savannas can be divided into three main types, the more open arid woodland, moist woodlands which are referred to as the Miombo and only occur north of South Africa, and Mopani woodlands that dominate many of the low lying and hot river valley systems (Olson et al. 2001) (Fig. 2.7). The Fynbos Biome and Succulent Karoo Biome have exceptionally high plant species diversity and endemism. Key features of these biomes and the climatic and disturbance envelopes in which they occur are described below.

Terrestrial biomes of southern Africa derived from Mucina and Rutherford (2006) for South Africa, merged with (Olson et al. 2001) for the rest of southern Africa. Olson’s ecoregions were reclassified to match the South African biomes. Halophytics (salt pans) and flooded grasslands (e.g., the Okavango) are distinctive features in the Savanna Biome and not true biomes

Typical appearances of southern African terrestrial biomes: (1) Desert, (2) Succulent-Karoo, (3) Indian-Ocean Coastal Belt, (4) Nama-Karoo, (5) Savanna, (6) Grassland, (7) Forest, (8) Albany Thicket, (9) Fynbos; 1–3 top, 4–6 middle, 7–9 bottom row. Photos 2, 5, 7–9 GvM. 1, 3, 4, 6 Dreamstime.com royalty free license
3.2.1 Savanna Biome
Savannas are by far the dominant biome of southern Africa covering 2.16 million km2 (65%) of southern Africa and additionally extending north to as far as the Sahel, covering 60% of Africa (Scholes and Walker 1993). The Savanna Biome is distinctive in that there is a codominance of both a woody component including both perennial trees and shrubs (phanaerophytes) and herbaceous layer (hemicryptophytes) of predominantly C4 grasses, but also many forbs. Embedded in the savanna are also halophytics pans such as Etosha in Namibia and the Makgadikgadi Pan in Botswana, in total about 0.8% of the southern African region. In addition, there are flooded grasslands, the Okavango delta being the largest and most famous at a further 0.8% of the area. Both of these unique habitats add to the savanna habitat diversity and biodiversity.
Southern African savanna occurs over a rainfall gradient from less than 250 mm yr−1 to about 1800 mm yr−1 (Scholes and Walker 1993). It is, however, limited to areas that have wet summers and long dry winters. In southern Africa savanna is found in areas of unimodal rainfall, while in east Africa, it occurs in areas of bimodal rainfall. Tree cover in the savanna ranges widely from under 1% to about 80%, and is normally below the ecological limits for maximum tree canopy cover (Sankaran et al. 2005; Chap. 15). Effects of fire, herbivory and climatic variability play an important role in maintaining the dynamics of savanna systems, especially in the moist areas which in the absence of these disturbances could become forests (Sankaran et al. 2005; Chap. 14). Fire is a common occurrence ranging from occasional at the dry extreme to almost annual in moist areas (Archibald et al. 2010). Herbivores, including the so-called mega herbivores and especially elephants, also appear to play a major role in the savanna dynamics (Bond et al. 2003; Stevens et al. 2016). The grass component of savanna is resistant to both grazing and fire impacts, mature trees tend to be fire resistant, but tree seedlings are often susceptible to fires. Crown fires are rare, and even if the crowns are destroyed, most species can resprout.
Southern African Savanna is part of the Sudano-Zambezian phytochorion (White 1983). A major distinction is between what is termed ‘moist savanna’ or Miombo in higher rainfall regions which has predominantly broad-leaf trees and a nutrient-poor substratum, as opposed to the ‘arid savanna’ or ‘bushveld’ in more arid regions with nutrient-rich substrates and predominantly fine-leafed trees, and finally the ‘mopane savanna’, which is similar to the arid savanna but dominated by the tree Colocospermum mopane (Huntley 1982; Chap. 15). Savannas have high alpha diversity (i.e., species richness at a local site) and contain 3–14 species per m2 and 40–100 species per 0.1 ha, but with low local endemism (Scholes and Walker 1993).
3.2.2 Grassland Biome
The Grassland Biome is the second largest biome in southern Africa (360,000 km2, 11%), and with the exception of small areas in the highlands of Zimbabwe, is almost exclusively limited to South Africa. A single stratum of hemicryptophytes is what characterizes the grasslands. Trees and shrubs are largely absent or limited to fire refugia. Five centers of plant endemism have been identified within the grasslands. The grasslands have a high flora diversity, especially of nongraminoid herbaceous plants with geophytes (perennial plants) being of particular importance. Grasslands have high alpha diversity and may contain 9–49 species per 100 m2 plot, with a 1000 m2 plot containing 55–100 species (Mucina and Rutherford 2006). C4 grasses dominate in the warmer and arid areas, while C3 grasses dominate at cooler high-altitude sites. The grasslands can be divided into two main classes, moist areas on leached and dystrophic soils that tend to be ‘sour’ with low palatability and the more arid grasses which are ‘sweet’, even when dormant, and palatable to livestock (Ellery 1992).
Grasslands occur over a wide range of rainfall from about 400 to 1200 mm annually, and over a wide range of soils, ranging from sea level to >3300 m altitude and located in areas that extend from frost free to snow covered in winter (O’Connor and Bredenkamp 2003). As in savannas, fire is an important driver of the ecology of grasslands, and is one of the main reasons that areas of high rainfall do not become forests. Small forest patches are often found within fire refugia in grasslands. There has been extensive debate as to why grasslands persist as grassland, despite the moister areas being able to support forest. Ellery (1992) suggested that fire and grazing regimes exclude woody plants. However, there appears to be a limit to either, as heavy grazing and altered fire regimes can change grass species composition and degrade the grassland. The grasslands are susceptible to invasions of alien plant species, including herbaceous species such as Pompom weed (Campuloclinium macrocephalum) and trees such as the Australian Acacia mearnsii (Black wattle), which may not only change the species composition but the entire structure. Invasion of indigenous woody species into grasslands is also an issue of concern, and this is a process similar to the bush encroachment experiences in savanna (Skowno et al. 2017; Chap. 15). The grasslands are one of the most anthropogenically transformed biomes as they are suitable for annual crops on the good soil flat areas and for timber plantations in the higher rainfall mountainous areas.
3.2.3 Nama-Karoo Biome
The Nama-Karoo Biome (260,000 km2; 8% of the area) is found in South Africa and Namibia at altitudes between 550 and 1500 m. It is an arid environment dominated by perennial dwarf shrubs and annual grass. The vegetation furthermore includes leaf succulents, stem succulents, bulbous monocotyledons and annuals (Marloth 1908). Gibbs Russell (1987) classified the Nama-Karoo Biome as being 50% hemicryptophytes, and 25% each of chamaephytes and cryptophytes. Gradients across the biome go from strongly dwarf-shrub dominated in the west, to a higher grass ratio in the east where the biome meets the grassland and savanna biomes. Rainfall occurs predominantly in summer and ranges from about 60 to 400 mm/yr. Rainfall is highly variable, particularly in the most arid regions. Although the soils tend to be heavy and eutrophic, they are typically very shallow, with calcareous layers being common (Palmer and Hoffman 2003).
3.2.4 Desert Biome
Desert (112,000 km2; 3% of the area) is found almost exclusively along the coast of Namibia, though a narrow band is found in South Africa in the Senqu (Orange) river valley. It is often separated into the sand deserts to the south and the gravel plains to north. It is dominated by ephemeral plants that respond rapidly to the rare and unpredictable rainfall. There are also some unique perennials such as the Welwitschia mirabilis as well as trees growing along river beds. The Desert Biome has extremely low rainfall and a mean of 20 mm yr−1 is misleading as to annual rainfall as there can be multiple years of zero rainfall followed by a single substantive rainfall event. Dense fog does, however, compensate in part for the low rainfall, with a number of species such as the Stenocara gracilipes beetle exhibiting unique adaptations to harvest the fog.
3.2.5 Succulent Karoo Biome
This biome (107,000 km2) is found along the West Coast of South Africa and southern Namibia. It is characterized by winter rainfall of between 50 and 250 mm, supplemented by dewfall and fog. The biome has the highest biodiversity for any area globally with comparable rainfall, with very high levels of endemism. It is characterized by leaf-succulents and deciduous leafed dwarf shrubs dominated by the families Aizoaceae (Mesembryanthemaceae), Asteraceae, Crassulaceae and Euphorbiaceae (Desmet 2007). In addition, there are many annuals that form the main component of the mass spring floral display for which the biome is well known.
The Succulent Karoo Biome occupies a narrow niche between the Fynbos and the Nama-Karoo Biomes, where the former is characterized by winter rainfall, and the latter by summer rainfall. The Succulent Karoo vegetation is uniquely adapted to areas of low winter rainfall. Esler et al. (2015) have shown that it is fire that prevents the succulent karoo species from establishing within the higher rainfall fynbos. Any increase in precipitation that allows the fynbos to expand into the karoo would therefore be detrimental to the succulent karoo vegetation. Equally, any shift in seasonality toward summer rainfall dominance would likely allow for Nama-Karoo species to expand into the Succulent Karoo Biome.
3.2.6 Fynbos Biome
The Fynbos Biome is a small biome of about 85,000 km2, only 2.6% of southern Africa and only occurs in South Africa, but has an extremely rich floral biodiversity of over 9000 vascular species (3% of the global vascular species) with 70% being endemic (Goldblatt and Manning 2002; Rebelo et al. 2006). It also has high faunal endemism with 55% of its 44 frog taxa, 84% of its 19 freshwater fish and 31% of its 234 butterflies being endemic (Critical Ecosystem Partnership Fund 2001). A distinguishing feature of the vegetation is codominance by perennial shrubs or small trees with sclerophyllous microphyllous (small-leaved) leaf characteristics, sedges and the grass like Restionaceae. True grasses (Poaceae) are very rare in the west, but slightly less rare in the east of the biome (Rutherford and Westfall 1986). The distinctive climatic feature of the region is the predominantly winterly precipitation, driven by the seasonal mid-latitude cyclonic systems that sweep across the region. During summer these fronts are mostly pushed further south and miss the Cape region. Precipitation varies greatly, partly due to the influence of the mountainous topography, with some areas being as low as 210 mm/yr, while the mountain tops are some of the wettest areas in southern Africa with Jonkershoek reaching about 3000 mm/yr. The biome can establish over a wide range of altitudes, soil and aspect features, all probably important in the supporting the vegetation (Cowling et al. 2003a).
The Fynbos Biome, together with the Succulent Karoo Biome, constitutes the Cape Floristic Kingdom, one of only six floristic kingdoms recognized globally, and the only one to be contained within a single country. The Fynbos Biome can be divided into two key vegetation complexes, the renosterveld vegetation found on the more fertile, clay and silt soils of the lowlands and fynbos which dominates the predominantly nutrient-poor mountainous regions of the Table Mountain Group. Atypical areas are also linked to heavier shale band soils and limestone inselbergs. Three families, the Proteaceae, Ericaceae and reed-like Restionaceae dominate the vegetation, with the Proteaceae and Ericaceae in particular showing an exceptionally high level of what appears to be relatively recent Neogene and Quaternary speciation (Cowling et al. 2003a).
The vegetation is fire dependent and requires regular burns for its persistence (van Wilgen et al. 1994; van Wilgen et al. 2010). Optimum fire frequency is considered to be between 10 and 15 years, with intensity of the fire effect largely dependent on the season in which the fire occurs. Although several species resprout after fire, the predominate regeneration strategy for most species is through reseeding, and a multitude of seeding strategies have evolved. Both pollination and in some cases, seeding is dependent upon complex insect plant interactions (Le Maitre and Midgley 1992). The Fynbos has proved to be exceptionally vulnerable to the invasion of alien plants and this is posing a major threat to the region. These aliens, in addition to displacing natural vegetation, can also impact on fire frequency and intensity, further impacting on indigenous biodiversity (Richardson et al. 1996).
3.2.7 Forest Biome
True closed canopy forests are uncommon in southern Africa. The Forest Biome is defined as dominated by a tree layer or stratum ‘phanaerophytes’ with 75% or more, overlapping crown cover and graminoids, if present, are rare (Bailey et al. 1999; von Maltitz et al. 2003). This differs substantially from the FAO definition of forests which would include much of southern Africa’s savanna. An estimated 66,000 km2—2% of the region—is forest, most of it occurring in Mozambique. This excludes a further 0.2% of the area being Mangrove forest (based on Olson et al. 2001; Mucina and Rutherford 2006). However, it is unclear how the Indian Ocean Coastal Belt Biome in South Africa (Mucina and Rutherford 2006) and the Forest Biome (Olson et al. 2001) should be differentiated within Mozambique. Forest within the region has two key origins, Afrotemperate forest occurring in the higher altitude mountains of Zimbabwe and South Africa, and extending to the southern Cape of South Africa where latitude compensates for altitude, and the more tropical forests extending down the coast from Mozambique. Unlike the other biomes where the biome forms a large block of consolidated land, the forests tend to be embedded in the other biomes, typically in fire refugia. They occur in areas of over 525 mm yr−1 in winter rainfall regions or over 725 mm yr−1 in summer rainfall regions.
The Forest Biome has high species diversity, with 1438 plant species recorded for just the South African component. On a per area basis forest species diversity (0.58 species per km2) is second only to the fynbos (Geldenhuys 1992). Despite the low density of animals in forests, 14% of South Africa’s bird and mammal taxa have been recorded from forests (Geldenhuys and MacDevette 1989).
3.2.8 Indian Ocean Coastal Belt Biome
The species rich 34,000 km2 Indian Ocean Coastal Belt (IOCB) Biome is a narrow belt of vegetation stretching along the KwaZulu Natal coast in South Africa and along the coast of Mozambique in a vegetation type, which Olson et al. (2001) refer to as Southern Swahili coastal forests and woodlands. It contains both savanna and grassland elements and also includes embedded forest patches. The vegetation is a mosaic of areas with a factually and structurally savanna characteristics and areas of grassland characteristics. Although Moll and White (1978) recognized the uniqueness of the region, others such as Huntley (1982) have considered it part of the Savanna Biome and Low and Rebelo (1996) considered it a mix of grassland and savanna species. Rutherford and Westfall (1994) did not recognize it as an independent biome based of their classification systems, though Rutherford et al. (2006) finally gave it biome states, this based more on the tropical affinity of the biome, rather than its functional aspects, with the grasslands being either azonal or secondary in nature. The IOCB has unique species richness as it is part of the larger the Pondoland and Maputaland center of endemism hotspot, which is second only to the fynbos for species richness and endemism in South Africa, and contains over 8200 species of plants (23.5% endemic), 540 species of birds 200 mammal species, 200 reptiles (14.4% endemic) and 72 amphibians (15.3% endemic) (Conservation International 2014).
3.2.9 Albany Thicket Biome
Albany thicket is found exclusively within South Africa and is a 31,000 km2 area representing 1% of southern Africa. It consists of a dense, almost impenetrable, tangle of short trees less than 3 m tall and with almost 100% canopy cover. An herbaceous understory may be present, though with minimal grass. It is in a transitional zone between winter and summer rainfall and is found predominantly on fertile soils. It is hot and dry, with unpredictable low rainfall of 200 to 950 mm yr−1 (Mucina and Rutherford 2006). The vegetation has affinities with the karoo, savanna and grassland vegetation, though Cowling et al. (2005) suggest it is an ancient biome which can be traced back to the Eocene. It has a high plant diversity of about 2000 species of which about 300 are endemic to the biome. The succulent nature of many of the species and the relative scarcity of grass means that fire is not a major feature of the natural biome, but becomes common when the biome is degraded and grass starts to dominate. The biome is exceptionally susceptible to degradation from overgrazing, and a large proportion of the biome is already partially or severely degraded.

(Top) Seasonality of rain expressed as the ratio of summer rain (Oct, Nov, Dec, Jan, Feb, Mar) to winter rain (Apr, May, Jun, Jul, Aug, Sep), and (Bottom) mean annual precipitation (based on WorldClim 1970 to 2000 data, https://www.worldclim.org/data/worldclim21.html)
3.3 Diversity of Southern African Terrestrial Biomes
3.3.1 Large-Scale Environmental Factors Shaping Terrestrial Biodiversity
The variability in mean annual precipitation (MAP), seasonality of rainfall and temperature gradients, coupled with fire and disturbances, are factors that have led to the exceptional natural diversity of faunal and floral species within the southern African environment (Cowling et al. 2003a). Africa’s southernmost tip, the area of the Cape Floristic Kingdom, has evolved under a Mediterranean climate of winter rainfall and extreme summer water deficiency (Cowling et al. 2003b). As is discussed by Rouault et al. in Chap. 6 and shown in Fig. 2.9, the balance of the region has all year, or in most cases strongly summer-dominated precipitation with long arid winters, the perfect conditions for the development of grasslands and savanna vegetation (Huntley 1982; Scholes and Archer 1997). There are rainfall gradients going from less than 20 yr−1 precipitation in the West to over 3000 mm yr−1 in some of the mountainous areas (Fig. 2.9b). In addition, there is a high level of interseasonal variability of rainfall, linked mostly to the occurrence of El Niño Southern Oscillation (ENSO) cycles. This means that it is the norm, rather than the exception, to have periods of either above average rainfall, followed by similar periods with below average precipitation or even severe drought (Chaps. 6 and 29). The standard error of MAP is negatively correlated with rainfall, i.e., the most arid areas tend to have the most severe droughts but can also have exceptionally wet years. Not only has the region’s unique fauna and flora evolved to deal with this variability, but it is probable that this variability is critical to maintaining the flora and fauna in its current state.
The oceans surrounding southern Africa play an important role in moderating coastal temperatures, the West Coast dominated by the cold Atlantic currents, while the East Coast is impacted by warm Indian Ocean tropical currents (Fig. 2.1; Chap. 6). The prevailing winds causing upwelling of cold water in the Benguela Current along the West Coast are largely responsible for the desert environments along the West Coast, but also contribute to the formation of fog banks that have led to unique adaptations in plants and animals who use this fog as their main source of moisture. The relatively high-altitude inland plateau is both cooler and experiences greater fluctuations in temperature. This area can experience severe frost in winter, an important determinant of the interface between grassland and savanna woodland (Ellery et al. 1991).
Fire has played a critical role in defining vegetation biomes, and the boundaries between biomes are in many instances defined by the occurrence or lack of fire (Bond et al. 2003; Esler et al. 2015). Forest patches within the grasslands are found in fire refugia (von Maltitz et al. 2003), the Fynbos ends and the Succulent Karoo starts when fire is no longer supported (Esler et al. 2015), frost kills the aboveground component of grass plants in the moist grasslands which leads to intense and frequent fires that prevent savanna expansion into the grasslands (Ellery et al. 1991). Within the grasslands, savanna and fynbos vegetation fire is critical for the biome maintenance and its frequency and intensity impacts on floristic structure. Other disturbances such as the above-mentioned rainfall variability, browsing and grazing pressures, including impacts from the mega-herbivores are also gaining recognition for their role in biome maintenance.
3.3.2 Small-Scale Environmental Gradients Within Terrestrial Biomes
Within each biome there is a huge diversity of vegetation types driven by variation in local microclimatic conditions and soil properties. Within biomes, it is common to find areas with distinctly different geologies giving rise to very different soil nutrient status, and hence supporting completely different flora. For instance, much of the Fynbos is dominated by the nutrient-poor sandstone mountains of the Cape Folded Mountain belt. These bring about highly leached acidic soils. However, within the region, there are also limestone intrusions with base-rich soils. Similarly, much of the savanna is on eroded dystrophic granitic soils, sandstone or in the case of the extensive Kalahari, deep alluvial sands. These have distinctly different vegetation from the finer textured and nutrient-rich soils derived from basalt or shales.
Some areas of southern Africa have a great range of topography incorporating both relatively flat lowlands as well as mountains regions. Altitudinal gradients, varying aspects, differences in geologies and soils as well as the barriers to plant migration have contributed to extensive within biome diversity. For instance, in the Fynbos, the north facing and more harsh and arid mountain slopes are dominated by Restioids, while the Proteaceae are more prevalent on the cooler and moister southward oriented slopes at the same altitudes (Cowling et al. 2003b).
At a finer scale, there are typically unique plant communities associated with different catena positions. The savannas in particular have well-developed catenas where the ridge crest tends to have deep sandy soils, the midslope has shallow soils, often with a thin hydromorphic grass-belt along seep-lines in situations where moisture is forced to the surface by a clay plug. Clay soils with comparatively high nutrient content are found in the valley bottoms (Gertenbach 1983). On the granite-derived soils in Kruger National Park, the vegetation at the top of the catena has moist savanna features with trees being broad leafed and the grass largely unpalatable. By contrast the valley bottom has arid savanna characteristics of palatable grass and microphylls trees (see also Chap. 15). Unique habitats such a rivers, sodic areas and saltpans add additional habitat diversity within biomes.
Disturbance regimes can also play an important role in creating different habitats and diversity of the associated flora and fauna. In this regard the return periodicity of fire is of particular importance in all the fire dependent biomes. Frequent fires in the fynbos can prevent many of the more woody species, many of which are obligatory reseeders, from reaching reproductive maturity. In the savanna, frequent fire tends to favor grass over trees. A mosaic of different fire histories therefore assists in maintaining high diversity, both at the level of plant functional types as well as species (Parr and Brocket 1999). At an even finer scale, grazing lawns created by herbivores congregating, have higher nutrient states and a differing species composition than the surrounding savanna (Hempson et al. 2015).
Other disturbances such as grazing or browsing pressures, including the extreme impacts from mega herbivores such as elephants, the occurrence of droughts and floods, interact with fire in maintaining diversity. There is a growing acceptance that many of the southern African biomes are best understood based on disequilibrium theory (Nakanyala et al. 2018).
3.3.3 Primary and Secondary Productivity and Use
Modeled gross primary production (GPP) and mean annual net primary production (NPP) largely reflect mean annual rainfall patterns, except for areas with winter- and all-year rainfall, where NPP is substantially lower than the amount of rainfall would suggest. Individual biomes differ widely in their range of annual production (Table 2.1). Field-based studies on NPP are few (see Scholes et al. 2014), though there are numerous satellite-based estimates where trends in NPP are estimated (see Chap. 26).
For all biomes, except for Forest, the principle land use of untransformed vegetation can be described as rangeland, leading to secondary production in the form of livestock. However, the fynbos vegetation tends to have a very low carrying capacity for livestock or wildlife (over 30 hectares per large stock unit (LSU) (based on a 450 kg steer) for the renosterveld and over 100 ha LSU−1 for mountain fynbos) due to low levels of palatability, partly as a consequence of the key characteristics of the sclerophyllous plants and the low nutrient status of the soils. The Succulent Karoo Biome and Desert Biomes have very low carrying capacity for livestock due to the low rainfall, this despite them mostly having eutrophic soils. The remaining biomes are extensively used either for livestock production or increasingly for wildlife (see Chaps. 16 and 18). The grasslands (4–6 ha LSU−1) and savanna (10–15 ha LSU−1) in particular are highly suited to livestock and game. Although primary productivity increases with rainfall, soils tend to become more leached (Huntley 1982). There tends to be a critical point above about 600 mm yr−1 of precipitation where soils become leached and dystrophic, causing the grass and trees to switch from being predominantly palatable in winter, to being unpalatable when dry in winter (Huntley 1982). Soil nutrient status, texture and catena position can alter the precipitation level where this change takes place. This gives rise to what is termed ‘sweetveld’ which can be grazed all year in the arid areas and ‘sourveld’ which is only palatable during the summer in the moister areas. The interplay between NPP, palatability and grazing pressure means that there is high build-up of unpalatable grassy biomass in the high rainfall areas, and this is a driver for frequent fires in these areas (see Chap. 26). The Albany Thicket Biome is a remarkable outlier. In its natural state it has a higher NPP than might be expected for its low rainfall, and can support a high livestock of wildlife density. This is in large part due to the spekboom (Portulacaria afra) which has a joint CAM and C3 photosynthetic pathway and is both highly palatable and relatively fast-growing. Thicket is very sensitive to overgrazing, particularly by goats. When overutilized the thicket vegetation can ‘collapse,’ changing from a productive thicket (dense stands of small tree)-based system to a low-production annual grassland, with a concomitant loss of soil and biomass. There is evidence that the thicket is more resilient to wildlife and particularly elephant grazing, compared to grazing by livestock, and it has been identified as an area where rewilding might have extensive carbon sequestration benefits (Mills et al. 2005; Mills and Cowling 2010).
The Nama-Karoo Biome, despite its relatively low productivity, once supported vast herds of migratory herbivores. The vegetation has a carrying capacity of about 25–35 ha LSU−1 (Cowling and Roux, 1987). Human settlement has caused the area to be fenced into camps and it now supports an important sheep and goat industry (Masubelele et al. 2015). There are long-standing concerns over the impacts of overgrazing on the overall degradation of the karoo vegetation and in particular changes in the shrubs to grasses ratios. Acocks (1953) already raised concerns over the karoo shrubs encroaching into the grasslands. More recent data suggest that the opposite might in fact be true and that climate change is causing grasses to increase in the karoo (Masubelele et al. 2014). See Chap. 17 in this volume for recent work on the differences in production from highly grazed versus lightly grazed karoo areas.
Both the grasslands and the renosterveld are areas targeted for agricultural crop production. This has led to extensive transformation of these vegetation types in areas where good soils and adequate rainfall overlap. The grasslands and to a lesser extent the mountain Fynbos areas are also targeted for plantation forestry production, again causing transformation of the natural vegetation.
Forests have some of the highest rates of NPP in the region. However, southern Africa has very limited areas of true, closed-canopy forests, and they mostly lie in relatively cool climates which lower NPP rates. Higher production rates are observed in the more tropical coastal forests or on the East Coast of South Africa and Mozambique, rather than in the cooler mountain and southern Cape forests. Most of the South African forest is under conservation, though a limited amount of legal, sustainable timber and forest product harvesting takes place. In addition, an unsanctioned harvesting for traditional uses including traditional medicines also takes place, though there is a recent trend toward joint forest management to better regulate these traditional uses (Geldenhuys 2004). In Mozambique forests are extensively harvested, much of this illegally for export, but also for local charcoal production and the opening of new agricultural fields (Nielsen and Bunkenborg 2020; Woollen et al. 2016). Plantation forests of exotic pine, eucalyptus and wattle (Acacias) provide most of the region’s timber, and pulp for paper, but represent a fully transformed, but productive habitat.
In addition to provision of grazing the natural vegetation provides humans with a multitude of what are termed ‘veld products’ or nontimber forest products. These products are especially important to the rural poor and vulnerable (Chap. 15). These products include medicinal plants for traditional remedies, especially from bark and geophytes, thatching grass, wild fruits, mushrooms and other foods, building material and craft material. Rural communities throughout southern Africa are also still dependent on fuelwood as the main source of domestic cooking fuel (Malimbwi et al. 2010). This dependency on wood fuel is partially reduced due to rural electrification, but even in electrified areas is still a common practice to use wood to spare costs. In Mozambique, charcoal is also the main urban domestic fuel (Mudombi et al. 2018) of the poor and there is an extensive charcoal trade between rural and urban centers (Smith et al. 2019). Within the Fynbos biome there is a large industry based on wild flower harvesting and the harvesting of rooibos tea (Aspalathus linearis), though both are now mostly grown in plantation format on transformed or semitransformed land (Louw 2006).
3.3.4 Organizational Efforts Geared to Protect Terrestrial Biodiversity
Southern Africa has a long and proud history of terrestrial biodiversity conservation. As a region it has, by global standards, both a high proportion of land under formal conservation and a high proportion of still largely natural environments. Historically, low human density, less destructive farming practices, unsuitability of land for year-round human habitation, and the occurrences of diseases such as malaria and trypanosomiasis (sleeping sickness) resulted in many areas remaining relatively natural. Western technology and medicine resulted in rapid population expansion in the twentieth century, coupled with opening of new land areas for agriculture, and this necessitated the need for formal conservation. Conservation effort is, however, not strategically allocated to conserving all biodiversity, and in many cases, it is not linked to the likely threats to the biodiversity of the biomes. The savanna is well conserved as in addition to large parts of the savanna biome being covered with formal conservation, there are also huge areas of private land under wildlife management, and extensive tracts of communal land in Namibia or the old hunting areas of Botswana which are managed by communities as part of Community-Based Natural Resource Management (CBNRM) tourism programs which adds to the conservation of the biome (see Chaps. 15 and 17).
Overall, the biodiversity-rich Fynbos Biome enjoys about 20% formal conservation; however, conservation differs significantly between the fynbos versus the renosterbos vegetation. Not only is the renosterbos poorly conserved at only 0.6%, but it has been extensively transformed for cropland (96% transformed). The conserved fynbos areas are predominantly in mountain regions, which are historically conserved as water catchments. Impacts from invasive alien vegetation occur throughout the biome and threaten the fynbos in both protected and unprotected areas. The Cape Action for People and the Environment (C.A.P.E.) (1998–2000), developed a 20-year strategy for conservation and sustainable development of the fynbos biodiversity hotspot. Funding from the Global Environment Facility (GEF), Critically Endanger Partnership Fund, National Government and numerous other donors has helped implement this plan.
Only about 3.5% of species- and endemic-rich Succulent Karoo Biome is formally conserved. The Succulent Karoo Ecosystem Plan (SKEP) was a participative process to improve conservation of the endangered Succulent Karoo biome, and created a 20-year strategic plan (Driver et al. 2003). It used a strategic conservation planning approach to identify critical areas needing conservation. Further funded through the GEF, Critical Ecosystems Partnership Fund, in collaboration with other funders added more than 2.9 million hectares of biologically important land to the conservation estate via establishment of conservancies, signing of stewardship agreements, and the incorporation and designation of state land. This program included both South African and Namibia (Critical Ecosystem Partnership Fund 2001).
The Grassland Biome is extensively transformed into cropland and plantation forestry, and despite being one of the most threatened biomes, only has 3.2% formal conservation. The Grasslands Program was initiated by SANBI to strengthen conservation within the grasslands (DEA 2015).
The Nama-Karoo Biome has exceptionally low levels of formal conservation in South Africa, with only about 1% of the biome conserved. Much of the South African Nama-Karoo is considered degraded from overgrazing, but total land transformation is relatively limited (Hoffman 2014). Compared to South Africa, conservation levels are better in Namibia where about 17% of the Nama-Karoo Biome is conserved.
The Desert Biome is well conserved in both South Africa and Namibia. The Forests Biome within South Africa has an exceptionally high level of conservation as even forests outside of formal conservation areas have special protection under the National Forest Act. None of the Mozambique forest is within formal reserves, though a small proportion is in a hunting area. In South Africa both the IOCB and Albany Thicket have below the international 10% target for conservation, and both are vulnerable to degradation and transformation. Within Mozambique a small section of the IOCR is protected.
Initial conservation efforts in the region tended to focus on land with a low agricultural potential, rather than the strategic importance of the land from a biodiversity conservation perspective. South Africa has an active program to increase the extent of conservation, and in addition many areas throughout the region have been identified for management through studentship agreement. Several transboundary conservation initiatives have been implemented in the region and there is strong NGO support through organizations such as the Peace Park Foundation and WWF. Although conservation areas are relatively well managed and successful throughout most of the southern African region, funding for conservation remains problematic in all countries. In Mozambique conservation areas largely collapsed during the civil war and management capacity was lost. To help fill this gap, the private sector has become involved supporting formal conservation in initiatives such as the Carr foundations support to Gorongosa reserve in Mozambique, and the Frankfort Zoological Society’s supports the Gonarezhou reserve in Zimbabwe. Ecotourism has also been used as a funding mechanism to support private and community conservation initiatives.
4 Connection Between Oceanic and Coastal Terrestrial Ecosystems
Land and ocean in the Benguela region of southern Africa are closely interlinked. The desert-dominated coastal strip along the West Coast is dependent on water and nutrient inputs via the sea fog, just as the coastal waters are influenced by nutrient inputs via desert dust and water inputs via submarine groundwater discharge. Climate change influences these material fluxes considerably (Bryant et al. 2007). The connections between these fluxes and ecological processes are not yet fully understood, and hence, the extent of this impact on pelagic and benthic ecosystems in the coastal waters off Namibia, on the Namib Desert and on the greenhouse-gas concentrations in the atmosphere has not yet been quantified.
The main transport vectors are winds, fog, river and groundwater discharges (Fig. 2.10). These vectors are driven by the complex interplay between the ocean and the atmospheric circulation over southern Africa and its teleconnections with climate anomalies reflected by indicators such as the El Niño Southern Oscillation (ENSO), Tropical Southern Atlantic Index (TSA) or St Helena Island Climate Index (HIX).

Sketch of the main environmental factors, transport processes, and ecosystem components in the Namibian coastal upwelling-desert system (© Werner Ekau)
The relatively cold sea surface in the upwelling system of the Benguela Current in combination with subtropical subsidence lead to a rather stable lower atmosphere, which suppresses the formation of substantial precipitation. Therefore, other sources of moisture are of tremendous importance in the region. Due to the cold waters of the Benguela Current, advected sea fog is a rather frequent feature (Schulze 1969; Lancaster et al. 1984) along the southern African coastal region and provides a major source of moisture for several local species (Hachfeld 2000; Hamilton III et al. 2003, Henschel and Seely 2008). Maximum monthly precipitation (dew) from fog can reach up to about 50 mm yr−1, while annual mean rainfall amounts (excluding fog precipitation) for the region are only between ~20 and 40 mm yr−1 (Lancaster et al. 1984).
Fog thus is a main source of water for the Namib Desert ecosystem. It occurs mainly during the austral winter months when extremely cold and nutrient-/CO2-rich water wells up along the Namibian coast (Eckardt and Schemenauer 1998). The generation of fog is enhanced when low atmospheric pressure systems develop in the south and upwelling is restricted to a narrow strip along the coast in the north. Under such circumstances warm moist air of the south and/or the open ocean is transported over upwelled water where it cools down. Sea surface temperatures have increased over the last 30 years and upwelling is assumed to strengthen in the course of global warming, raising the question of how this might affect the fog formation over the Namib Desert.
In the land-sea direction, another connecting water transport exists through submarine groundwater discharge (SGD). In Namibia for instance, the two most important coastal aquifers, the Omaruru Delta and the Kuiseb Dune, are closely linked to paleoriver channels filled with sands which provide the permeability for subsurface flow toward the sea. Submarine groundwater can have significant impact on shallow-water benthic communities (Starke et al. 2019).

Dust plumes along the Namibian coast (https://eoimages.gsfc.nasa.gov)
Besides water, fog contains salt and nutrients which indicate that sea water is not its only source. It appears to act as a barrier scavenging all sorts of dust and other atmospheric compounds. When the water evaporates, salt crusts are formed within the coastal deserts (Eckardt and Schemenauer 1998). Fog is acidic compared to sea water, but contains higher concentrations of dissolved inorganic carbon and calcium carbonate than rain water. Several desert plants such as lichens and grasses, which stabilize the soil in the central Namib, are dependent upon the moisture and nutrients released from the fog.
Dust inputs into the ocean are mainly controlled by the bergwinds channeled through the ephemeral river beds (see Fig. 2.11). Besides inorganic dust, winds transport also organic particles such as desert-grass, lichens and insects providing additional food for the marine and the desert fauna and the associated food webs (Fig. 2.9). Barkley et al. (2019) recently showed that apart from dust, the aerosol transport of African biomass burning is a substantial source of phosphorus deposition to the Amazon Basin, the tropical Atlantic and the Southern Ocean.
Sediment traps which have been deployed in the framework of SPACES along the Namibian continental margin between the Walvis Ridge and the Orange River are, in turn, tools to measure the flux of particles from the surface into the deep ocean. Sediment trap results show that the majority of the exported material is residue from the pelagic food web but contains on average also ~9% to 17% of inorganic dust particles referred to as lithogenic matter (Vorrath et al. 2018). A long-term sediment trap deployment close to Walvis Bay on the Namibian shelf (see Chap. 25 for more detailed information) showed furthermore that the contribution of lithogenic dust varies significantly on time scales of weeks but also seasonally and interannually. For instance, at the end of 2010, lithogenic dust particles contributed more than 40% to the trapped material whereas at the end of the year 2013 the contribution of lithogenic matter to total flux was less than 10%. These dust inputs are known to act as ballast in sinking particles which lowers the offshore advection of plankton blooms, and as essential sources of micronutrients such as iron which is assumed to limit plankton blooms in many other ocean regions (Rixen et al. 2019, and references therein). To what extent the varying dust inputs affect the development of plankton blooms still needs to be studied in more detail to better understand impacts of climate and local land use changes on the marine environment.
5 Southern African Biomes: Carbon Sources or Sinks?
5.1 Oceanic Biomes
Numerical models suggest that upwelling systems at lower latitudes such as the Benguela Upwelling System (BUS) in the south-eastern Atlantic Ocean act as net carbon dioxide (CO2) sources to the atmosphere (Laruelle et al. 2014; Brady et al. 2019; Roobaert et al. 2019). In contrast, estimates based on data obtained during the SPACES program suggest that the nBUS acts as a CO2 source while the sBUS acts as a CO2 sink (Emeis et al. 2018; Siddiqui et al. 2023). These opposing functions are assumed to be a consequence of the combined effects of the marine solubility pump and the biological carbon pump on the partial pressure of CO2 (pCO2) in surface waters: if the pCO2 in the ocean falls below pCO2 in the atmosphere, the ocean takes up CO2, whereas it emits CO2 if the pCO2 in surface waters exceeds that in the atmosphere. The solubility pump increases pCO2 in seawater and favors CO2 emission when upwelled water warms at the surface. Conversely, the biological carbon pump reduces the pCO2 as it is driven by the photosynthesis of phytoplankton converting CO2 into biomass which is subsequently exported into subsurface waters. TRAFFIC research has supported the conclusion that the sBUS operates as a regional CO2 sink where the CO2 uptake by the biological carbon pump exceeds the CO2 release by the solubility pump, whereas the nBUS acts as a CO2 source to the atmosphere (Siddiqui et al. 2023).
However, effects of these pumps in the BUS have to be linked to their opposing function in the Southern Ocean, since the water that upwells in the BUS originates from there. According to our current estimate, CO2 uptake by the biological carbon pump in the BUS compensates up to 38% of its CO2 loss in the Atlantic sector of the Southern Ocean (Chap. 25). Hence, changes in CO2 uptake by the biological carbon pump in the BUS could significantly affect the marine carbon cycle and its role as a sink of anthropogenic CO2 (Siddiqui et al. 2023).
Even though it is widely accepted that global change (Riebesell et al. 2007) and human impacts via, e.g., fisheries (Bianchi et al. 2021), affect the biological carbon pump, associated changes of its CO2 uptake are still difficult to predict due to the low accuracy with which it can be determined. Accordingly, the development of new methods and observing strategies remain a great challenge in marine sciences, and efforts to protect the oceans and combat climate change need to be strengthened (Chap. 25).
5.2 Terrestrial Biomes
A 2014 South African assessment of terrestrial carbon stocks, repeated again in 2020 found that to a large extent standing carbon stocks follow rainfall patterns. Grasslands have one of the highest carbon stocks per unit area, in some cases approaching those of forests (Scholes et al. 2014; von Maltitz et al. 2020). This un-intuitive outcome is because of the extensive soil carbon stocks that accumulate particularly in the moist high-altitude grasslands. The savanna, despite having a slightly lower carbon stock per unit area, is a globally important carbon stock due to the vastness of the savanna regions.
Carbon fluxes remain poorly understood for the region and are covered in greater detail in subsequent chapters (Chaps. 17 and 30). A number of factors make understanding of the carbon fluxes complicated. Firstly, there are the overarching impacts from global climate change and the global fertilization impacts from raised CO2 on standing plant biomass. According to Knowles et al. (2014), this should have a positive impact on most, but not all, biomes. Secondly, for most of the biomes, there is a gradual uptake of carbon over multiple years. This follows seasonal patterns of uptake during the wet periods, with emissions during the dry periods. However, in the fire-prone biomes, there are also major emissions during fire periods that might be every year in some grasslands to once in 20 years or more in some of the arid savanna and fynbos habitats. Especially in the fynbos, this leads to an almost total mortality of standing woody biomass, followed by reseeding and a gradual build-up of new biomass. Long-term trends are therefore difficult to ascertain, especially since fire intervals can vary greatly on the same parcel of land over time. Thirdly, the region has cycles of wet and arid periods that can span many years (Chaps. 3 and 6). This can mean that short-term rainfall-induced trends are very different from longer-term climate change and CO2 fertilization trends. Further, most satellite-derived biomass products are only starting to give a sufficient time span of data to overrule the short-term rainfall fluctuations. Finally, human induced land degradation and restoration need to be considered (von Maltitz et al. 2018). Even using long-term data from National Parks can give misleading results. For instance, in the Kruger National Park elephants were almost absent when the park was proclaimed in 1902. Elephant numbers increased and were artificially held constant from 1967 to 1994 at about 7000 to 8000 animals; however, since 1994, their numbers have been allowed to increase and now stand at about 18,000. This has had profound impacts on standing tree biomass, as can be seen at the flux tower near Skukuza which has changed from a relatively dense savanna (30% tree cover) when initiated to a more open savanna at present (Scholes et al. 2001).
6 Impacts of Climate Change
Anthropogenic climate change is projected to affect individual terrestrial and marine biomes to different degrees. In marine ecosystems, regional atmospheric warming interacts with altered ocean currents. In terrestrial systems, aside from direct effects of warming, altered rainfall causes the main effects on biomes under climate change. Shifts in tropical cyclone behavior are not yet projected with reasonable confidence, although they may fundamentally change the extent and steepness of environmental gradients that have existed for millennia.
6.1 Impact of Climate Change on Marine Biomes
6.1.1 Climate Change Affecting Coastal Upwelling Systems
All Eastern Boundary Upwelling Systems (EBUSs) are dependent on the global circulation patterns of wind and ocean currents and thus are susceptible to alterations caused under climate change. Climate change affects EBUSs in different ways with partly opposing consequences. Global warming of the ocean surface will increase stratification and, hence, potentially reduce upwelling intensity. In contrast, increasing greenhouse gas concentrations in the atmosphere could force intensification of upwelling-favorable winds in EBUSs (stronger trade winds) and, therefore, strengthen upwelling intensity (Bakun et al. 2015). Thus, both opposing trends, the intensification and the relaxation of upwelling intensity, are possible under climate change scenarios. It is not yet clear which of the two processes will dominate where, but latitudinal differences in EBUSs’ response to global climate change appear likely. Recent studies suggest that a poleward shift in subtropical high-pressure cells will lead to an intensification, and possibly seasonal expansion, of upwelling in poleward parts of EBUSs, whereas closer to the equator, upwelling intensity will be reduced (García-Reyes et al. 2015; Rykaczewski et al. 2015; Wang et al. 2015). Some evidence for this phenomenon has been observed in the Benguela System, including a significant recent decrease in upwelling in the nBUS, and a significant increase on the Agulhas Bank off South Africa (Lamont et al. 2018).
Sweijd and Smit (2020) showed that 99% of the area of all seven African Large Marine Ecosystems (LMEs) has warmed with rates of between 0.11°C per decade (Agulhas LME) and 0.58°C per decade (Canary LME) since the 1980s. They found that only 1% of the LME area was associated with cooling due to upwelling. The most intense warming hotspot in the nBUS was at the Angola-Benguela Frontal Zone, while the strongest cooling was at the boundary between the nBUS and sBUS, in the Lüderitz upwelling cell.
Sydeman et al. (2014) evaluated the evidence for an intensification of upwelling-favorable winds through a meta-analysis of existing studies and found support for the hypothesis that climate change is associated with stronger coastal winds and enhanced upwelling in EBUSs, including the Benguela. This in turn could lead to adverse effects such as stronger offshore advection, more frequent hypoxic events, and increased ocean acidification (Bakun et al. 2015). Recent studies within the SPACES framework point out the extreme complexity of oceanographic processes on a smaller scale (e.g., eddies), which may also have modulating effects on the wind-driven upwelling off Lüderitz and Walvis Bay (Bordbar et al. 2021).
The Benguela Current Ecosystem is vulnerable to processes leading to deoxygenation. Increasing temperatures and decreasing oxygen levels have been measured—especially in the nBUS—for several decades (Stramma et al. 2008) with severe potential impact on the species there (Ekau et al. 2010). Deoxygenation is especially pronounced in the nBUS, as this subsystem is already defined by perennial low-oxygen water and hydrogen sulfide eruptions (Kirkman et al. 2016).
Whereas a variety of studies have shown that the impacts of global warming (and other anthropogenic stressors, e.g., fishing pressure or plastic pollution) on the marine food web and ecosystem health vary spatially, even within the Benguela Upwelling System, the region-specific effects in many areas of southern Africa have not been well studied (Hutchings et al. 2012; Kirkman et al. 2016; Verheye et al. 2016).
6.1.2 Climate Change Impacts on Marine Species in the Benguela
Climate change is expected to have complex effects with potentially far-reaching consequences in the vulnerable Benguela Upwelling System (Bakun et al. 2015; Bordbar et al. 2021). The contrasting local warming and cooling trends may have opposing effects on individual taxa and will cause complex ecosystem responses to climate change.
Phytoplankton production would principally be favored by enhanced coastal upwelling leading to elevated nutrient concentrations in surface waters, but it could simultaneously be reduced by greater turbulence and thus deeper mixed layers and light limitation (Bakun et al. 2015). Increased turbulence and offshore export may also cause a decoupling between primary producers and higher trophic levels (spatial mismatch). In contrast, new primary production further offshore may be enhanced. Alterations in phytoplankton and zooplankton communities will likely be the consequence, but are difficult to predict at the current state of knowledge. Certainly, such changes will propagate up the food web, e.g., impacting larval fish recruitment.
The expansion of oxygen-minimum zones (Stramma et al. 2008) affects the vertical distribution and migration of zoo- and ichthyoplankton (Ekau and Verheye 2005; Auel and Ekau 2009) that respond to the shift in oxyclines. Here, as in the Humboldt and California Current EBUSs (Bograd et al. 2008; Bertrand et al. 2010), the position of the oxycline is a determining factor in the structuring of the pelagic community as it limits the habitable space for pelagic organisms (Howard et al. 2020). In the zooplankton, certain species e.g., copepods of the families Eucalanidae, Rhincalanidae and Subeucalanidae, can better cope with oxygen minimum zones (OMZs) than others (Teuber et al. 2013; Teuber et al. 2019). The shifting of the oxycline in the nBUS could have severe impacts on fish recruitment due to the high sensitivity of many fish species to low-oxygen concentrations, and also via changing availability of zooplankton prey (Kreiner et al. 2009; Ekau et al. 2018). Overall, deoxygenation may have multiple ecological consequences, including range shifts in species distribution or alteration of vertical migration behavior, which in turn affect species interactions, as well as the local abundance and availability of commercially relevant living marine resources (Chan et al. 2019).
Potts et al. (2015) reviewed projected impacts of climate change (sea temperature, upwelling intensity, current strength, rainfall, pH, and sea level) on coastal fishes in southern Africa, distinguishing the effects on different groups of fishes (migratory, resident, estuarine-dependent or catadromous fish guilds) in separate biogeographical zones. The authors concluded that impacts will be diverse due to the different life strategies of coastal fishes, but process understanding would still need to be improved in order to derive sound predictions for the different zones of the southern African coasts.
In the northern Benguela, long-term trends in growth of fish stocks such as the deep-water hake were shown to be influenced by upwelling intensity, as well as by fisheries-induced evolution, which works in opposing directions for small and large fish (Wilhelm et al. 2020). Therefore, for each fished and non-fished taxon, understanding of their specific responses to the combined climate change and fishing impacts still needs to be improved.
For key species in the southern Benguela, Ortega-Cisneros et al. (2018a) performed a trait-based sensitivity assessment in order to evaluate their potential relative sensitivity to climate change. Several of the species included in their assessment are endemic to southern Africa, including white steenbras Lithognathus lithognathus, which was classified as particularly sensitive. The benthic invertebrate abalone (Haliotis midae) also has high expected vulnerability. Cape horse mackerel, shallow-water hake and sardine scored as having lower sensitivity to climate change than the other assessed species. Nevertheless, sardine, in particular, appears to suffer from the combined effects of climate change and fishing.
The South African fishery on small pelagics targets sardine, anchovy and (to a lesser degree) round herring, and is economically the country’s second most valuable fishery (after the deep-sea trawl fisheries) and supports many employees. These characteristics, together with the fact that small pelagic fishes respond rapidly to environmental change in terms of their abundance and distribution patterns, have led to it being considered one of the fisheries most vulnerable to climate change (van der Lingen 2021). The sardine resource has been low for more than a decade, and is presently depleted, while both anchovy and sardine have shown changes in their distributions. Various adaptation measures have been suggested and/or implemented to mitigate climate change effects and to nurse the pelagic stocks. These range from sardine stock rebuilding programs and the importation of frozen sardines to reduce local fishing pressure while maintaining the processing infrastructure, to developing experimental fisheries on alternative resources, specifically the mesopelagic Hector’s lanternfish (Lampanyctodes hectoris). Augustyn et al. (2018) present various rebuilding measures for the sardine fishery and point out that monitoring and scientific data collection are vital to continue recording the most important environmental variables and species to enable adaptive management of that fishery.
The complexity of interactions of partly antagonistic climate-driven processes makes overall predictions difficult, and strongly suggests that climate impact will differ spatially. The frequency of occurrence of harmful algal blooms has been suggested to increase under climate change, and negative effects on sardine (but not on anchovy or round herring) have been described (van der Lingen et al. 2016). In contrast to the other small pelagic species, sardine possesses finely meshed gill rakers which enable them to feed directly on small unicellular phytoplankton, but also making them vulnerable to the possible ingestion of toxic dinoflagellates such as Gonyaulax polygramma directly (van der Lingen et al. 2016). As sardine and anchovy differ in feeding behavior and diet, changes in local zooplankton community composition and size structure can also be expected to affect each fish species differently (van der Lingen et al. 2006a, b). Where larger zooplankton become more abundant, anchovy and round herring may be favored, whereas sardine may gain a competitive advantage where smaller zooplankton will dominate particularly under conditions of weak upwelling (van der Lingen et al. 2006b; Augustyn et al. 2018).
Physical mesoscale processes which directly impact the habitat of small pelagic fishes cause considerable ‘noise’ in oceanographic data, making detection of climate change signals difficult (van der Lingen and Hampton 2018). This complicates efforts to disentangle effects of climate change and fishing activities. Therefore, an integrated research response needs to be developed in order to improve forecasting of likely climate change impacts on vulnerable fish species and the related fishing sectors (van der Lingen 2021).
In a modeling study investigating the effects of several drivers on the southern Benguela upwelling system, Ortega-Cisneros et al. (2018b) found that warming had the greatest effect on species biomass, with mainly negative effects reported. Cephalopods are an exception to this rule, as there have been various records of their thriving under climate change (Golikov et al. 2013; Doubleday et al. 2016; Oesterwind et al. 2022). Mesopelagic fishes and large pelagic fish species were predicted to be the least and the most negatively affected groups, respectively, also when accounting for the effects of potential new fisheries on mesopelagics and round herring (Ortega-Cisneros et al. 2018b). Van der Sleen et al. (2022) incorporated sea surface temperature (SST) fluctuations in models of marine fish population dynamics at different trophic levels in ten LMEs (including the Agulhas Current LME). Using observed landings and SST data, they constructed their model to disentangle influences of climate from those of fishing pressure with some predictive power. Yet, in upwelling systems, responses to SST are partly opposing and generally more complex.
6.2 Impact of Climate Change on Terrestrial Biomes
Climate change is anticipated to have major impacts on the terrestrial biomes. A number of research initiatives starting from simple bioclimatic envelope approaches (Midgley et al. 2008) and progressing to complex Dynamic Vegetation Models (Chap. 14). Outcomes from the modeling processes differ based on the different approaches, and the different climate futures used to drive the models, though shrinkage of many of the biomes appears inevitable. Climate change can be expected to affect the individual terrestrial biomes differently.
Savannas are a relatively resilient vegetation type as they have evolved under a multitude of disturbances; however, it is possible that climate change may move savannas beyond critical tipping points. Direct impacts from higher temperature are unlikely to affect savannas, though the increased temperatures may make the area more arid (DEA 2015). However, climate change poses a number of potential new threats to the biome. The C4 grasses evolved relatively recently under low global CO2 concentrations (Bouchenak-Khelladi et al. 2009). As climate change is associated with elevated levels of atmospheric CO2, this may lead to a shift in the competitive advantage of entire metabolic classes of plants. The C4 photosynthetic pathway currently gives the C4 grasses a water use efficiency advantage. Rising CO2 levels appear, however, to favor C3 species, and indications are that this will favor trees over the grasses, possibly shifting the biome to higher tree density or even forest (Bond et al. 2003; Kgope et al. 2010; Chap. 14). Fire regimes under climate change remain a key unknown. Increased tree density might reduce fire risks; however, there is also a possibility that prolonged droughts and heat waves under climate change may lead to unprecedented ‘fire storms’ with unknown consequence to the tree-grass ratios (Staver et al. 2011).
Grasslands are considered as especially vulnerable to climate change, with models suggesting they will be encroached by other biomes and ‘squeezed’ to a fraction of their current extent (e.g., Midgley et al. 2008; DEA 2015; Chap. 14). Tree species are expanding into grasslands on the savanna grassland interface, effecting millions of ha of grasslands (Skowno et al. 2017). Grasslands may, however, expand into the Karoo in some circumstances (Chap. 15), and their greater prevalence in some Nama-Karoo shrublands may already be driving a novel wildfire regime.
The Nama-Karoo Biome, while being typified by the occurrence of dwarf shrubs of the Asteraceae, Poaceae, Aizoaceae, Mesembryanthemacaea, Liliaceae and Scrophulariaceae, may also have a large amount of grass, especially following periods of high rainfall (Palmer and Hoffman 2003). The aridity index of the biome has changed over the past 100 years and there is an increase in grass cover since the 1960s (Hoffman 2014). Fire is rare in the Nama-Karoo Biome, and although most of the Nama-Karoo shrubs are able to resprout after fire, fire could shift the vegetation to a more grassy state (Kraaij et al. 2017). Morgan et al. (2011) suggest that increased CO2 concentrations may favor C4 grasses, potentially increasing the likelihood of fire which will further benefit grass more than shrubs (du Toit 2019). Projected climate change impacts on the Nama-Karoo vegetation are unclear, especially when utilizing projections of both climate and geology, but may reduce the Nama-Karoo Biome in South Africa, though cause its expansion in Namibia and Botswana (Guo et al. 2017).
The fauna and flora of the Desert Biome are critically dependent on the moisture provided through fog. This biome is considered exceptionally vulnerable to changes in fog occurrence, which is poorly understood, but considered to be declining under climate change (Mitchell et al. 2020).
For the Succulent Karoo Biome, any increase in precipitation that allows the fynbos species to expand would be detrimental. At the same time, any shift in seasonality toward stronger summer rainfall would likely allow for Nama-Karoo species expansion at the expense of the succulent karoo species. As such the Succulent Karoo Biome is considered as extremely vulnerable to small shifts in climate. Early biome niche models predict that the biome might shrink considerably as a result of a southward shift in the rain-bearing mid latitude cyclones (MacKellar et al. 2007; Midgley and Thuiller 2007). A model by Driver et al. (2012) based on more recent climate predictions found lesser evidence of extensive biome shrinkage.
The renosterveld element of the Fynbos Biome in particular has been highly fragmented through wide-scale transformation. The region is considered as very vulnerable to climate change impacts, due to the intrinsic nature of its high, but localized and isolated biodiversity. Increased temperature regimes could change fire frequency and intensity, factors that are known to impact fynbos biodiversity. Being at the southern tip of Africa, only altitude, and not southward migration, is available as an option for a range shift to avoid increased heat. A southerly shift of only a few 10s of km of the rainfall-bearing frontal systems that provide the unique winter rainfall could be devastating to the region.
Forests are sensitive to fire, and there are records of forests burning during intense hot and dry periods when the resultant damage can be catastrophic (Geldenhuys 1994). This makes them potentially vulnerable to climate change impacts where droughts and heat waves may become both more common and more intense.
Being close to the warm Indian Ocean Current, climate is subtropical in the Indian Ocean Coastal Belt Biome, but extremes are moderated by the coastal influence. Rainfall tends to be above 900 mm yr−1. Rainfall occurs predominantly during summer in the south, but throughout the year toward the north. Climate change will have a lesser impact on temperature in the IOCB than in the inland biomes, and the east of the country where it occurs also appears to be slightly less vulnerable to an overall decrease in rainfall. Changes in fire regimes due to climate change might be one of the greatest climate change induced threats (DEA 2015).
Climate change impacts on the Albany Thicket Biome are poorly understood, but a predicted increase in rainfall might favor an incursion of savanna species including more grass into the biome. The Crassulacean Acid Metabolism (CAM) dominance of many of the species is less likely to benefit from raised global CO2 which will favor C3 trees and to a lesser degree also C4 grasses. Increased occurrence of fire in this biome would be devastating to some species (DEA 2015).
The Desert Biome is projected to expand from its current distribution in coastal and southern Namibia and western southern Africa, toward the south and east, displacing Nama Karoo and savanna in western Botswana and northern South Africa. The iconic desert tree Aloidendron dichotomum may already be showing signs of population mortality in warmer and drier areas of its range in these regions (Foden et al. 2007).
7 Conclusions and Implications
The southern African subcontinent and its surrounding oceanic regions accommodate globally unique ecoregions, which are outstanding owing to their exceptional biodiversity, with the occurrence of a multitude of endemic species and, in the marine realm, a very high productivity. Southern African terrestrial and marine ecosystems are shaped by steep and extended environmental gradients; they are adapted to and dependent on these special conditions. The biota in terrestrial biomes are limited in potential adaptive range extensions toward the south. Therefore, the effects of climate change on specific biomes will call for changes in human use of resources on land and at sea, and for protective measures to safeguard the exceptionally high biodiversity of the subcontinent and its waters. In marine ecosystems, the effects of climate change are complex, especially since the underlying mechanisms are partly counteractive, particularly in the Benguela Upwelling System: global warming leads to stronger thermal stratification of the water column, impacting many biological processes (e.g., time of spawning, distribution) and biogeochemical vertical exchange. Simultaneously, higher gradients in air pressure and temperature changes over land and ocean also alter the regional wind fields. This may increase turbulence in the pelagic realm and intensify the upwelling of nutrient-rich waters, providing resources for pelagic food webs in some regions, or reduce upwelling in other regions. At the same time, various additional anthropogenic stressors including high fishing pressure are bound to threaten the ecological balance in marine ecosystems and their services such as the sequestration of CO2. These processes may exacerbate the effects of climate change currently predicted.
Southern Africa, especially South Africa, has accumulated a long history of extensive biological research, unsurpassed by most areas outside of Europe and going back to the seventeenth century (Huntley 2003). These efforts have generated unique terrestrial and oceanic data sets of high value both for Africa and for most of the rest of the developing world. Data sets are stored in both European and local databases (e.g., PANGAEA, ODINAFRICA, ocean.gov.za) and involve numerous world-class national universities and research institutes. Many of these databases are constantly being expanded, becoming even more valuable. Their availability in extensive publications and ongoing studies on biodiversity and ecosystem functioning provide one of the most comprehensive settings for research investment both for regional and global benefit.
Current regional and international research regarding the biomes of southern Africa does not focus on the biology alone, but also addresses earth system processes and particularly climate change. Southern Africa has also generated extensive cross-disciplinary research interests beyond the continent, leading to numerous long-term collaborative research initiatives with Europe and other high-income nations. This includes programs such as Biodiversity Monitoring Transect Analysis (BIOTA) Africa, the Southern African Science Service Centre for Climate Change and Adaptive Land Management (SASSCAL), and Science Partnerships for the Adaptation to Complex Earth System Processes in Southern Africa (SPACES) I and II programs focusing on the region. South Africa is also a driving partner in the All Atlantic Ocean Research and Innovation Alliance, so far including Atlantic coastal countries Brazil, South Africa, Argentina, Cabo Verde, Morocco, United States, Canada and several European states, and supported by the European Commission (relating to Galway Statement, Belém Statement and Washington Declaration; https://allatlanticocean.org/). These large initiatives increase the knowledge base needed to enable decisions toward sustainable management of human activities.
Their outstanding value, and at the same time their great vulnerability through climate change, habitat loss and various other human pressures, should make southern African biodiversity the focus of future research activities aimed at providing scientific support for ecosystem conservation measures and their management. The present digital age provides an excellent opportunity to create databases, making data sets readily available to simplify scientific and political collaboration and to exchange and thereby formulate socially, environmentally and economically acceptable management aims. However, various prerequisites need to be considered: the rich historic data sets need to be kept relevant by ensuring ongoing monitoring across all biota. This is a basic requirement for the analyses, meta-analyses and evaluations performed by scientists of different disciplines who need to formulate regular status reports on ecosystem health (e.g., status report on fisheries in South Africa, DEFF 2020) either for each country, or preferably through transnational organizations and institutional collaborations. These status reports could be evaluated by interdisciplinary committees to formulate up-to-date and clearly defined socio-economic management aims and guidelines, which at the same time protect the southern African biodiversity as well as ecosystem health and services. These steps advance society toward the global Sustainable Development Goals of finding socio-economic balance hand in hand with ecological balances.
Notes
- 1.
TRAFFIC - Trophic Transfer Efficiency in the Benguela Current
- 2.
SPACES II – Science Partnerships for the Adaptation to Complex Earth System Processes in Southern Africa
References
Acocks JPH (1953) Veld types of South Africa. Memoirs of the Botanical Surveys South Africa, vol 28, Government Printer, South Africa, 192 p
Archibald S, Nickless A, Govender N, Scholes RJ, Lehsten V (2010) Climate and the inter-annual variability of fire in southern Africa: a meta-analysis using long-term field data and satellite-derived burnt area data. Glob Ecol Biogeogr 19:794–809
Auel H, Ekau W (2009) Distribution and respiration of the high-latitude pelagic amphipod Themisto gaudichaudii in the Benguela Current in relation to upwelling intensity. Prog Oceanogr 83:237–241
Augustyn CJ, Cockcroft A, Coetzee J, Durholtz D, van der Lingen CD (2018) Rebuilding South African fisheries: three case-studies. In: Garcia SM, Ye Y (eds) Rebuilding of Marine Fisheries part 2: case studies, Fisheries and Aquaculture Technical Paper No. 630/2. FAO, Rome
Awad AA, Griffiths CL, Turpie JK (2002) Distribution of South African marine benthic invertebrates applied to the selection of priority conservation areas. Divers Distrib 8:129–145
Ayón P, Swartzman G, Bertrand A, Gutiérrez M, Bertrand S (2008) Zooplankton and forage fish species off Peru: large-scale bottom-up forcing and local-scale depletion. Prog Oceanogr 79:208–214
Bailey C, Shackleton C, Geldenhuys C, Moshe D, Flemming G, Vink E, Rathogwa N, Cawe S (1999) Guide to and summary of the meta-database pertaining to selected attributes of South African indigenous forests and woodlands. In: Division of Water EaFT, CSIR (ed) Report ENV-P-C 99027, Pretoria
Bakun A, Black BA, Bograd SJ, Garcia-Reyes M, Miller AJ, Rykaczewski RR, Sydeman WJ (2015) Anticipated effects of climate change on coastal upwelling ecosystems. Curr Clim Change Rep 1:85–93
Barange M, Stuart V (1991) Distribution patterns, abundance and population dynamics of the euphausiids Nyctiphanes capensis and Euphausia hanseni in the northern Benguela upwelling system. Mar Biol 109:93–101
Barkley AE, Prospero JM, Mahowald N, Hamilton DS, Popendorf KJ, Oehlert AM, Pourmand A, Gatineau A, Panechou-Pulcherie K, Blackwelder P, Gaston CJ (2019) African biomass burning is a substantial source of phosphorus deposition to the Amazon, tropical Atlantic Ocean, and Southern Ocean. Proc Natl Acad Sci 116:16216–16221
Batten SD, Abu-Alhaija R, Chiba S, Edwards M, Graham G, Jyothibabu R, Kitchener JA, Koubbi P, McQuatters-Gollop A, Muxagata E, Ostle C, Richardson A, Robinson K, Takahashi K, Verheye H, Wilson W (2019) A global plankton diversity monitoring program. Front Mar Sci 6:321
BCC (2018) Regional strategy for marine spatial planning in the Benguela Current Large Marine Ecosystem. Benguela Current Convention, Swakopmund
Beal LM, De Ruijter WP, Biastoch A, Zahn R, SCOR/WCRP/IAPSO Working Group 136 (2011) On the role of the Agulhas System in ocean circulation and climate. Nature 472:429–436
Bertrand A, Ballon M, Chaigneau A (2010) Acoustic observation of living organisms reveals the upper limit of the oxygen minimum zone. PLoS One 5:e10330
Bianchi D, Carozza David A, Galbraith Eric D, Guiet J, DeVries T (2021) Estimating global biomass and biogeochemical cycling of marine fish with and without fishing. Sci Adv 7:eabd7554
Biggs R, Scholes R (2002) Land-cover changes in South Africa 1911-1993: research in action. S Afr J Sci 98:420–424
Bode M, Kreiner A, van der Plas AK, Louw DC, Horaeb R, Auel H, Hagen W (2014) Spatio-temporal variability of copepod abundance along the 20°S monitoring transect in the northern Benguela upwelling system from 2005 to 2011. PLoS One 9:e97738
Bode M, Hagen W, Cornils A, Kaiser P, Auel H (2018) Copepod distribution and biodiversity patterns from the surface to the deep sea along a latitudinal transect in the eastern Atlantic Ocean (24°N to 21°S). Prog Oceanogr 161:66–77
Bograd SJ, Castro CG, Di Lorenzo E, Palacios DM, Bailey H, Gilly W, Chavez FP (2008) Oxygen declines and the shoaling of the hypoxic boundary in the California Current. Geophys Res Lett 35:1–6
Bohata K (2015) Microzooplankton of the northern Benguela upwelling system. PhD Thesis, Staats- und Universitätsbibliothek Hamburg, 163 pp
Bond W, Midgley G, Woodward F (2003) What controls South African vegetation - climate or fire? S Afr J Bot 69:79–91
Bond WJ, Woodward FI, Midgley GF (2005) The global distribution of ecosystems in a world without fire. New Phytol 165:525–538
Bordbar MH, Mohrholz V, Schmidt M (2021) The relation of wind-driven coastal and offshore upwelling in the Benguela Upwelling System. J Phys Oceanogr 51:3117–3133
Bouchenak-Khelladi Y, Anthony Verboom G, Hodkinson TR, Salamin N, Francois O, Ni Chonghaile G, Savolainen V (2009) The origins and diversification of C4 grasses and savanna-adapted ungulates. Glob Chang Biol 15:2397–2417
Boyer D, Oelofsen B (2004) Co-management: Namibia’s experience with two large-scale industrial fisheries – sardine and orange roughy. In: Sumaila UR, Boyer D, Skogen MD, Steinshamn SI (eds) Namibia’s fisheries: Ecological, Economic and Social Aspects. Eburon Academic Publishers, pp 333–356
Brady RX, Lovenduski NS, Alexander MA, Jacox M, Gruber N (2019) On the role of climate modes in modulating the air–sea CO2 fluxes in eastern boundary upwelling systems. Biogeosciences 16:329–346
Branch WR (2006) A plan for phylogenetic studies of southern African reptiles. In: Branch WR, Tolley KA, Cunningham M, Bauer AM, Alexander G, Harrison JA, Turner AA, Bates MF (eds) SANBI biodiversity series. A plan for phylogenetic studies of southern African reptiles: proceedings of a workshop held at Kirstenbosch, February 2006. South African National Biodiversity Institute (SANBI)
Branch WR (2014) Conservation status, diversity, endemism, hotspots and threats. In: Bates MF, Branch WR, Bauer AM, Burger M, Marais J, Alexander GJ, de Villiers MS (eds) Atlas and red list of the reptiles of South Africa, Lesotho and Swaziland. Suricata Series, vol 1. South African National Biodiversity Institute (SANBI), Pretoria
Briggs JC, Bowen BW (2012) A realignment of marine biogeographic provinces with particular reference to fish distributions. J Biogeogr 39:12–30
Brodeur RD, Link JS, Smith BE, Ford M, Kobayashi D, Jones TT (2016) Ecological and economic consequences of ignoring jellyfish: a plea for increased monitoring of ecosystems. Fisheries 41:630–637
Brüchert V, Currie B, Peard KR, Lass U, Endler R, Dübecke A, Julies E, Leipe T, Zitzmann S (2006) Biogeochemical and physical control on shelf anoxia and water column hydrogen sulphide in the Benguela coastal upwelling system off Namibia. In: Neretin LN (ed) Past and present water column anoxia. Springer, Dordrecht
Bryant RG, Bigg GR, Mahowald NM, Eckardt FD, Ross SG (2007) Dust emission response to climate in southern Africa. J Geophys Res-Atmos 112:D09207, https://doi:10.1029/2005JD007025
Chan F, Barth JA, Kroeker KJ, Lubchenco J, Menge BA (2019) The dynamics and impact of ocean acidification and hypoxia - insights from sustained investigations in the northern California Current Large Marine Ecosystem. Oceanography 32:62–71
Chavez FP, Messié M (2009) A comparison of eastern boundary upwelling ecosystems. Prog Oceanogr 83:80–96
Coetzee J, Staby A, Krakstad J-O, Stenevik E (2009) Abundance and distribution of mesopelagic fish in the Benguela ecosystem, pp 44–47. In: Hampton I, Barange M, Sweijd N (eds). Benguela Environment Fisheries Interaction and Training Programme (BENEFIT) research projects. GLOBEC Report 25: ii, 126 p
Coetzee J, Merkle D, Shabangu F, Geja Y, Petersen J (2018) Results of the 2018 pelagic biomass survey. Fisheries/2018/DEC/SWG-PEL, vol 38. Department of Agriculture, Forestry and Fisheries, Cape Town
Conservation International (2014) Maputaland-Pondoland-Albany. Copy viewed on 17 March 2014. http://www.conservation.org/where/priority_areas/hotspots/africa/Maputaland-Pondoland-Albany/Pages/default.aspx
Costello MJ, Coll M, Danovaro R, Halpin P, Ojaveer H, Miloslavich P (2010) A census of marine biodiversity knowledge, resources, and future challenges. PLoS One 5:e12110
Cowling R, Roux P (eds) (1987) Karoo biome: a preliminary synthesis. Part 2-vegetation and history, vol 142. National Scientific Programmes Unit: CSIR
Cowling RM, Richardson DM, Mustart P (2003a) Fynbos. In: Cowling RM, Richardson DM, Pierce SM (eds) Vegetation of southern Africa. Cambridge University Press, Cambridge
Cowling RM, Richardson DM, Schulze RE, Hoffman MT, Midgley JJ, Hilton-Taylor C (2003b) Species diversity at the regional scale. In: Cowling RM, Richardson DM, Pierce SM (eds) Vegetation of southern Africa. Cambridge University Press, Cambridge
Cowling R, Procheş Ş, Vlok J, Van Staden J (2005) On the origin of southern African subtropical thicket vegetation. S Afr J Bot 71:1–23
Cowling RM, Potts AJ, Franklin J, Midgley GF, Engelbrecht F, Marean CW (2020) Describing a drowned Pleistocene ecosystem: last glacial maximum vegetation reconstruction of the Palaeo-Agulhas plain. Quat Sci Rev 235:105866
Crawford RJM, Underhill LG, Coetzee JC, Fairweather T, Shannon LJ, Wolfaardt AC (2008) Influences of the abundance and distribution of prey on African penguins Spheniscus demersus off western South Africa. Afr J Mar Sci 30:167–175
Critical Ecosystem Partnership Fund (2001) Ecosystem profile. The Cape floristic region, South Africa. https://www.cepf.net/resources/ecosystem-profile-documents/cape-floristic-region-ecosystem-profile-2001
Currie B, Utne-Palm AC, Salvanes AGV (2018) Winning ways with hydrogen sulphide on the Namibian shelf. Front Mar Sci 5:341
Cury P, Roy C (1989) Optimal environmental window and pelagic fish recruitment success in upwelling areas. Can J Fish Aquat Sci 46:670–680
Cury P, Bakun A, Crawford RJ, Jarre A, Quinones RA, Shannon LJ, Verheye HM (2000) Small pelagics in upwelling systems: patterns of interaction and structural changes in ‘wasp-waist’ ecosystems. ICES J Mar Sci 57:603–618
Cushing DH (1996) Towards a science of recruitment in fish populations, vol 7. Ecology Institute Oldendorf/Luhe Nordbünte
De Decker AHB (1964) Observations on the ecology and distribution of Copepoda in the marine plankton of South Africa. In: Investigational report of the Division of Sea Fisheries, vol 49. Department of Commerce and Industries, South Africa
De Decker A (1984) Near-surface copepod distribution in the south-western Indian and south-eastern Atlantic Ocean. Ann S Afr Mus 93:303–370
De Decker A, Kaczmaruk B, Marska G (1991) A new species of Calanus (Copepoda, Calanoida) from South African waters. Ann S Afr Mus 101:27–44
DEA (2015) Climate change adaptation plans for South African biomes. In: Kharika JRM, Mkhize NCS, Munyai T, Khavhagali VP, Davis C, Dziba D, Scholes R, van Garderen E, von Maltitz G, Le Maitre D, Archibald S, Lotter D, van Deventer H, Midgely G, Hoffman T (eds) Department of Environmental Affairs. Pretoria, South Africa
DEFF (2020) Status of the South African marine fisheries resources 2020. Department of Environment, Forestry and Fisheries, Cape Town, South Africa
Desmet P (2007) Namaqualand—a brief overview of the physical and floristic environment. J Arid Environ 70:570–587
Dey SP, Vichi M, Fearon G, Seyboth E, Findlay KP, Meynecke J-O, de Bie J, Lee SB, Samanta S, Barraqueta JLM, Roychoudhury AN, Mackey B (2021) Oceanographic anomalies coinciding with humpback whale super-group occurrences in the southern Benguela. Sci Rep 11:1–13
Dippenaar-Schoeman AS, Lyle R, Marais P, Haddad C, Foord S, Lotz L (2015) South African National Survey of Arachnida (SANSA): review of current knowledge, constraints and future needs for documenting spider diversity (Arachnida: Araneae). Trans R Soc South Africa 70:245–275
Doubleday ZA, Prowse TA, Arkhipkin A, Pierce GJ, Semmens J, Steer M, Leporati SC, Lourenço S, Quetglas A, Sauer W (2016) Global proliferation of cephalopods. Curr Biol 26:R406–R407
Driver A, Desmet P, Rouget M, Cowling R, Maze K (2003) Succulent Karoo ecosystem plan: biodiversity component. Technical Report CCU 1/03. Cape Conservation Unit, Botanical Society of South Africa
Driver A, Sink KJ, Nel JL, Holness S, Van Niekerk L, Daniels F, Jonas Z, Majiedt PA, Harris L, Maze K (2012) National Biodiversity Assessment 2011: an assessment of South Africa’s biodiversity and ecosystems. Synthesis report. South African National Biodiversity Institute (SANBI) and Department of Environmental Affairs, Pretoria, South Africa
du Toit JCO (2019) Drivers of vegetation change in the eastern Karoo. PhD thesis, University of KwaZulu-Natal, Pietermaritzburg, South Africa, pp 183
Duncan SE, Sell AF, Hagen W, Fock HO (2022) Environmental drivers of upper mesopelagic fish assemblages in the Benguela Upwelling Systems. Mar Ecol Prog Ser 688:133–152
Duncombe Rae CM (2005) A demonstration of the hydrographic partition of the Benguela upwelling ecosystem at 26°40'S. Afr J Mar Sci 27:617–628
Durholtz M, Singh L, Fairweather T, Leslie R, van der Lingen C, Bross C, Hutchings L, Rademeyer R, Butterworth D, Payne A (2015) Fisheries, ecology and markets of South African hake. In: Arancibia H (ed) Hakes: biology and exploitation. John Wiley & Sons, Chichester
Eckardt FD, Schemenauer RS (1998) Fog water chemistry in the Namib Desert, Namibia. Atmos Environ 32:2595–2599
Ekau W, Verheye HM (2005) Influence of oceanographic fronts and low oxygen on the distribution of ichthyoplankton in the Benguela and southern Angola currents. Afr J Mar Sci 27:629–639
Ekau W, Hendricks A, Kadler S, Koch V, Loick N (2001) Winter ichthyoplankton in the northern Benguela upwelling and Angola-Benguela front regions: BENEFIT marine science. S Afr J Sci 97:259–265
Ekau W, Auel H, Pörtner H-O, Gilbert D (2010) Impacts of hypoxia on the structure and processes in pelagic communities (zooplankton, macro-invertebrates and fish). Biogeosciences 7:1669–1699
Ekau W, Auel H, Hagen W, Koppelmann R, Wasmund N, Bohata K, Buchholz F, Geist S, Martin B, Schukat A, Verheye HM, Werner T (2018) Pelagic key species and mechanisms driving energy flows in the northern Benguela upwelling ecosystem and their feedback into biogeochemical cycles. J Mar Syst 188:49–62
Ellery WN (1992) Classification of vegetation of the South African grassland biome. PhD thesis, University of the Witwatersrand, Johannesburg, South Africa, pp 211
Ellery W, Scholes R, Mentis M (1991) An initial approach to predicting the sensitivity of the South African grassland biome to climate change. S Afr J Sci 87:499–503
Emeis K-C, Brüchert V, Currie B, Endler R, Ferdelman T, Kiessling A, Leipe T, Noli-Peard K, Struck U, Vogt T (2004) Shallow gas in shelf sediments of the Namibian coastal upwelling ecosystem. Cont Shelf Res 24:627–642
Emeis K, Eggert A, Flohr A, Lahajnar N, Nausch G, Neumann A, Rixen T, Schmidt M, van der Plas A, Wasmund N (2018) Biogeochemical processes and turnover rates in the northern Benguela upwelling system. J Mar Syst 188:63–80
Enquist BJ, Feng X, Boyle B, Maitner B, Newman EA, Jørgensen PM, Roehrdanz PR, Thiers BM, Burger JR, Corlett RT, Couvreur TLP, Dauby G, Donoghue JC, Foden W, Lovett JC, Marquet PA, Merow C, Midgley G, Morueta-Holme N, Neves DM, Oliveira-Filho AT, Kraft NJB, Park DS, Peet RK, Pillet M, Serra-Diaz JM, Sandel B, Schildhauer M, Simova I, Violle C, Wieringa JJ, Wiser SK, Hannah L, Svenning JC, McGill BJ (2019) The commonness of rarity: global and future distribution of rarity across land plants. Sci. Adv. 5:eaaz0414
Erasmus VN, Currie B, Roux J-P, Elwen SH, Kalola M, Tjizoo B, Kathena JN, Iitembu JA (2021) Predatory species left stranded following the collapse of the sardine Sardinops sagax (Pappe, 1854) stock off the northern Benguela upwelling system: a review. J Mar Syst 224:103623
Esler K, Von Staden L, Midgley G (2015) Determinants of the fynbos/succulent Karoo biome boundary: insights from a reciprocal transplant experiment. S Afr J Bot 101:120–128
Esteban I, Marean CW, Cowling RM, Fisher EC, Cabanes D, Albert RM (2020) Palaeoenvironments and plant availability during MIS 6 to MIS 3 on the edge of the Palaeo-Agulhas plain (South Coast, South Africa) as indicated by phytolith analysis at Pinnacle Point. Quat Sci Rev 235:105667
Fay A, McKinley G (2014) Global open-ocean biomes: mean and temporal variability. Earth System Sci Data 6:273–284
Finke G, Gee K, Gxaba T, Sorgenfrei R, Russo V, Pinto D, Nsiangango SE, Sousa LN, Braby R, Alves FL, Heinrichs B, Kreiner A, Amunyela M, Popose G, Ramakulukusha M, Naidoo A, Mausolf E, Nsingi KK (2020) Marine spatial planning in the Benguela Current Large Marine Ecosystem. Environ Dev 36:100569
Flynn RF, Granger J, Veitch JA, Siedlecki S, Burger JM, Pillay K, Fawcett SE (2020) On-shelf nutrient trapping enhances the fertility of the southern Benguela upwelling system. J Geophys Res Oceans 125:e2019JC015948
Foden W, Midgley GF, Hughes G, Bond WJ, Thuiller W, Hoffman MT, Kaleme P, Underhill LG, Rebelo A, Hannah L (2007) A changing climate is eroding the geographical range of the Namib Desert tree aloe through population declines and dispersal lags. Divers Distrib 13:645–653
Fortes-Lima C, Mtetwa E, Schlebusch C (eds) (2022) Africa, the cradle of human diversity: cultural and biological approaches to uncover African diversity. Brill, Leiden
García-Reyes M, Sydeman WJ, Schoeman DS, Rykaczewski RR, Black BA, Smit AJ and Bograd SJ (2015) Under Pressure: Climate Change, Upwelling, and Eastern Boundary Upwelling Ecosystems. Front. Mar. Sci. 2:109. https://doi.org/10.3389/fmars.2015.00109
Geldenhuys CJ (1992) Richness, composition and relationships of the floras of selected forests in southern Africa. Bothalia 22: 205–233 https://doi.org/10.4102/abc.v22i2.847
Geldenhuys CJ (1994) Bergwind fires and the location pattern of forest patches in the southern Cape landscape, South Africa. J Biogeogr 21:49–62
Geldenhuys CJ (2004) Bark harvesting for traditional medicine: from illegal resource degradation to participatory management. Scand J For Res 19:103–115
Geldenhuys C, MacDevette D (1989) Conservation status of coastal and montane evergreen forest. In: Huntley BJ (ed) Biotic diversity in southern Africa: concepts and conservation. Oxford University Press, Cape Town
Gertenbach WD (1983) Landscapes of the Kruger National Park. Koedoe 26:9–121
Gibbons MJ, Hutchings L (1996) Zooplankton diversity and community structure around southern Africa, with special attention to the Benguela upwelling system. S Afr J Sci 92:63–75
Gibbons MJ, Haddock SH, Matsumoto GI, Foster C (2021) Records of ctenophores from South Africa. PeerJ 9:e10697
Gibbs Russell G (1987) Preliminary floristic analysis of the major biomes in southern Africa. Bothalia 17:213–227
Goldblatt P, Manning JC (2002) Plant diversity of the Cape region of southern Africa. Ann Mo Bot Gard 89:281–302
Golikov AV, Sabirov RM, Lubin PA, Jørgensen LL (2013) Changes in distribution and range structure of Arctic cephalopods due to climatic changes of the last decades. Biodiversity 14:28–35
Good S, Fiedler E, Mao C, Martin MJ, Maycock A, Reid R, Roberts-Jones J, Searle T, Waters J, While J, Worsfold M (2020) The current configuration of the OSTIA system for operational production of foundation sea surface temperature and ice concentration analyses. Remote Sens 12:720
Griffiths CL (2005) Coastal marine biodiversity in East Africa. Indian J Mar Sci 34:35–41
Griffiths CL, Robinson TB (2016) Use and usefulness of measures of marine endemicity in South Africa. S Afr J Sci 112:1–7
Griffiths CL, Robinson TB, Lange L, Mead A (2010) Marine biodiversity in South Africa: an evaluation of current states of knowledge. PLoS One 5:e12008
Grote B, Ekau W, Hagen W, Huggett J, Verheye H (2007) Early life-history strategy of Cape hake in the Benguela upwelling region. Fish Res 86:179–187
Grote B, Stenevik E, Ekau W, Verheye H, Lipiński M, Hagen W (2012) Spawning strategies and transport of early stages of the two Cape hake species, Merluccius paradoxus and M. capensis, in the southern Benguela upwelling system. Afr J Mar Sci 34:195–204
Guo D, Desmet PG, Powrie LW (2017) Impact of the future changing climate on the southern Africa biomes, and the importance of geology. J Geosci Environ Prot 5:1–9
Hachfeld B (2000) Rain, fog and species richness in the central Namib Desert in the exceptional rainy season of 1999/2000. Dinteria 26:113–146
Hamilton WJ III, Henschel JR, Seely MK (2003) Fog collection by Namib Desert beetles: correspondence. S Afr J Sci 99:181
Hardman-Mountford NJ, Hirata T, Richardson KA, Aiken J (2008) An objective methodology for the classification of ecological pattern into biomes and provinces for the pelagic ocean. Remote Sens Environ 112:3341–3352
Harris LR, Holness SD, Finke G, Amunyela M, Braby R, Coelho N, Gee K, Kirkman SP, Kreiner A, Mausolf E, Majiedt P, Maletzky E, Nsingi KK, Russo V, Sink KJ and Sorgenfrei R (2022) Practical marine spatial management of ecologically or biologically significant marine areas: emerging lessons from evidence-based planning and implementation in a developing-world context. Front Mar Sci 9:831678. https://doi.org/10.3389/fmars.2022.831678
Hempson GP, Archibald S, Bond WJ, Ellis RP, Grant CC, Kruger FJ, Kruger LM, Moxley C, Owen-Smith N, Peel MJ, Smit IP, Vickers KJ (2015) Ecology of grazing lawns in Africa. Biol Rev 90:979–994
Henschel JR, Seely MK (2008) Ecophysiology of atmospheric moisture in the Namib Desert. Atmos Res 87:362–368
Heymans JJ, Tomczak MT (2016) Regime shifts in the northern Benguela ecosystem: challenges for management. Ecol Model 331:151–159
Hine P, Sealy J, Halkett D, Hart T (2010) Antiquity of stone-walled tidal fish traps on the Cape coast, South Africa. South Afr Archaeol Bull 65(191):35–44
Hoffman MT (2014) Changing patterns of rural land use and land cover in South Africa and their implications for land reform. J South Afr Stud 40:707–725
Howard EM, Penn JL, Frenzel H, Seibel BA, Bianchi D, Renault L, Kessouri F, Sutula MA, McWilliams JC, Deutsch C (2020) Climate-driven aerobic habitat loss in the California Current System. Sci Adv 6:eaay3188
Huggett J, Richardson A (2000) A review of the biology and ecology of Calanus agulhensis off South Africa. ICES J Mar Sci 57:1834–1849
Huggett J, Verheye H, Escribano R, Fairweather T (2009) Copepod biomass, size composition and production in the southern Benguela: Spatio–temporal patterns of variation, and comparison with other eastern boundary upwelling systems. Prog Oceanogr 83:197–207
Huggett JA, Noyon M, Carstensen J, Walker DR (2023) Patterns in the plankton – spatial distribution and long-term variability of copepods on the Agulhas Bank. Deep-Sea Res Part II 196:105265
Hugo S, Altwegg R (2017) The second southern African bird atlas project: causes and consequences of geographical sampling bias. Ecol Evol 7:6839–6849
Hulley PA, Lutjeharms JRE (1989) Lanternfishes of the southern Benguela region part 3. The pseudoceanic-oceanic interface. Ann S Afr Mus 98:1–10
Hulley P, Prosch R (1987) Mesopelagic fish derivatives in the southern Benguela upwelling region. S Afr J Mar Sci 5:597–611
Huntley B (1982) Southern African savannas. In: Huntley B, Walker B (eds) Ecology of tropical savannas. Springer, Berlin
Huntley B (2003) Introduction. In: Cowling RM, Richardson DM, Pierce SM (eds) Vegetation of southern Africa. Cambridge University Press, Cambridge
Huntley BJ, Russo V, Lages F, Ferrand N (2019) Biodiversity of Angola: science & conservation: a modern synthesis. Springer Nature, Cham
Hutchings L, Pillar SC, Verheye HM (1991) Estimates of standing stock, production and consumption of meso- and macrozooplankton in the Benguela ecosystem. S Afr J Mar Sci 11:499–512
Hutchings L, Beckley LE, Griffiths MH, Roberts MJ, Sundby S, van der Lingen C (2002) Spawning on the edge: spawning grounds and nursery areas around the southern African coastline. Mar Freshw Res 53:307–318
Hutchings L, van der Lingen CD, Shannon LJ, Crawford RJM, Verheye HMS, Bartholomae CH, van der Plas AK, Louw D, Kreiner A, Ostrowski M, Fidel Q, Barlow RG, Lamont T, Coetzee J, Shillington F, Veitch J, Currie JC, Monteiro PMS (2009) The Benguela Current: an ecosystem of four components. Prog Oceanogr 83:15–32
Hutchings L, Jarre A, Lamont T, Van den Berg M, Kirkman S (2012) St Helena Bay (southern Benguela) then and now: muted climate signals, large human impact. Afr J Mar Sci 34:559–583
IUCN (2016) A global standard for the identification of key biodiversity areas. Version 1.0. IUCN, Gland, Switzerland and Cambridge, UK
Jarre A, Ragaller SM, Hutchings L (2013) Long-term, ecosystem-scale changes in the southern Benguela marine pelagic social-ecological system: interaction of natural and human drivers. Ecol Soc 18(4):55
Jarre A, Hutchings L, Kirkman SP, Kreiner A, Tchipalanga PC, Kainge P, Uanivi U, van der Plas AK, Blamey LK, Coetzee JC, Lamont T, Samaai T, Verheye HM, Yemane DG, Axelsen BE, Ostrowski M, Stenevik EK, Loeng H (2015) Synthesis: climate effects on biodiversity, abundance and distribution of marine organisms in the Benguela. Fish Oceanogr 24:122–149
Kainge P, Kirkman SP, Estevão V, van der Lingen CD, Uanivi U, Kathena JN, van der Plas A, Githaiga-Mwicigi J, Makhado A, Nghimwatya L, Endjambi T, Paulus S, Kalola M, Antonio M, Tjizoo B, Shikongo T, Nsiangango S, Uahengo T, Bartholomae C, Mqoqi M, Hamukuaya H (2020) Fisheries yields, climate change, and ecosystem-based management of the Benguela Current Large Marine Ecosystem. Environ Dev 36:100567
Kgope BS, Bond WJ, Midgley GF (2010) Growth responses of African savanna trees implicate atmospheric [CO2] as a driver of past and current changes in savanna tree cover. Austral Ecol 35:451–463
Kirkman SP, Nsingi KK (2019) Marine biodiversity of Angola: biogeography and conservation. In: Huntley BJ, Russo V, Lages F, Ferrand N (eds) Biodiversity of Angola. Springer Open, Cham 43–52
Kirkman SP, Blamey L, Lamont T, Field JG, Bianchi G, Huggett JA, Hutchings L, Jackson-Veitch J, Jarre A, Lett C, Lipiński MR, Mafwila SW, Pfaff MC, Samaai T, Shannon LJ, Shin Y-J, van der Lingen CD, Yemane D (2016) Spatial characterisation of the Benguela ecosystem for ecosystem-based management. Afr J Mar Sci 38:7–22
Kirkman SP, Holness S, Harris LR, Sink KJ, Lombard AT, Kainge P, Majiedt P, Nsiangango SE, Nsingi KK, Samaai T (2019) Using systematic conservation planning to support marine spatial planning and achieve marine protection targets in the transboundary Benguela ecosystem. Ocean Coast Manag 168:117–129
Klak C, Reeves G, Hedderson T (2004) Unmatched tempo of evolution in southern African semi-desert ice plants. Nature 427:63–65
Knowles T, Boardman P, Mugido W, Blignaut J (2014) Understanding potential climate change mitigation opportunities. National Terrestrial Carbon Sinks Assessment: Technical Report. Department of Environmental Affairs, Pretoria, South Africa
Kopte R, Brandt P, Claus M, Greatbatch RJ, Dengler M (2018) Role of equatorial basin-mode resonance for the seasonal variability of the Angola Current at 11°S. J Phys Oceanogr 48:261–281
Kosobokova KN, Hopcroft RR, Hirche H-J (2011) Patterns of zooplankton diversity through the depths of the Arctic’s central basins. Mar Biodivers 41:29–50
Kraaij T, Young C, Bezuidenhout H (2017) Growth-form responses to fire in Nama-Karoo escarpment grassland, South Africa. Fire Ecol 13:85–94
Kreiner A, Stenevik EK, Ekau W (2009) Sardine Sardinops sagax and anchovy Engraulis encrasicolus larvae avoid regions with low dissolved oxygen concentration in the northern Benguela Current system J Fish Biol 74:270–277
Lamont T, García-Reyes M, Bograd S, van der Lingen C, Sydeman W (2018) Upwelling indices for comparative ecosystem studies: variability in the Benguela Upwelling System. J Mar Syst 188:3–16
Lancaster J, Lancaster N, Seely MK (1984) Climate of the central Namib Desert. Modoqua 14:5–61
Laruelle GG, Lauerwald R, Pfeil B, Regnier P (2014) Regionalized global budget of the CO2 exchange at the air-water interface in continental shelf seas. Glob Biogeochem Cycles 28:1199–1214
Le Maitre D, Midgley J (1992) Plant reproductive ecology. In: Cowling R (ed) The ecology of fynbos: nutrients, fire and diversity. Oxford University Press, Oxford
Lombard AT, Strauss T, Harris J, Sink K, Attwood C, Hutchings L (2004) South African national spatial biodiversity assessment 2004. Technical Report, vol 4: Marine Component. South African National Biodiversity Institute, Pretoria
Longhurst AR (2007) Ecological Geography of the Sea. Academic Press, Amsterdam
Louw R (2006) Sustainable harvesting of wild rooibos (Aspalathus linearis) in the Suid Bokkeveld, Northern Cape. MSc thesis, University of Cape Town, pp 130
Low A, Rebelo A (eds) (1996) Vegetation of South Africa, Lesotho and Swaziland. Department of Environmental Affairs and Tourism (DEAT), Pretoria, South Africa
Lynam CP, Gibbons MJ, Axelsen BE, Sparks CAJ, Coetzee J, Heywood BG, Brierley AS (2006) Jellyfish overtake fish in a heavily fished ecosystem. Curr Biol 16:R492–R493
MacKellar N, Hewitson B, Tadross M (2007) Namaqualand's climate: recent historical changes and future scenarios. J Arid Environ 70:604–614
Malimbwi R, Chidumayo E, Zahabu E, Kingazi S, Misana S, Luoga E, Nduwamungu J (2010) Woodfuel. In: Chidumayo E, Gumbo D (eds) The dry forests and woodlands of Africa managing for products and services. Earthscan, London
Marloth R (1908) Das Kapland. Gustav Fischer, Jena
Masubelele ML, Hoffman M, Bond W, Gambiza J (2014) A 50 year study shows grass cover has increased in shrublands of semi-arid South Africa. J Arid Environ 104:43–51
Masubelele M, Hoffman M, Bond W (2015) Biome stability and long-term vegetation change in the semi-arid, south-eastern interior of South Africa: a synthesis of repeat photo-monitoring studies. S Afr J Bot 101:139–147
MET (2018) Sixth National Report to the convention on biological diversity (2014–2018). Ministry of Environment and Tourism, Republic of Namibia, Windhoek, Namibia
Midgley G, Thuiller W (2007) Potential vulnerability of Namaqualand plant diversity to anthropogenic climate change. J Arid Environ 70:615–628
Midgley G, Rutherford M, Bond WJ (2008) The heat is on: impacts of climate change on plant diversity in South Africa. South African National Biodiversity Institute (SANBI), Pretoria
Mills AJ, Cowling RM (2010) Below-ground carbon stocks in intact and transformed subtropical thicket landscapes in semi-arid South Africa. J Arid Environ 74:93–100
Mills A, Cowling R, Fey M, Kerley G, Donaldson J, Lechmere-Oertel R, Sigwela A, Skowno A, Rundel P (2005) Effects of goat pastoralism on ecosystem carbon storage in semiarid thicket, eastern Cape, South Africa. Austral Ecol 30:797–804
Mitchell D, Henschel JR, Hetem RS, Wassenaar TD, Strauss WM, Hanrahan SA, Seely MK (2020) Fog and fauna of the Namib Desert: past and future. Ecosphere 11:e02996
Mohrholz V, Bartholomae CH, van der Plas AK, Lass HU (2008) The seasonal variability of the northern Benguela undercurrent and its relation to the oxygen budget on the shelf. Cont Shelf Res 28:424–441
Moll E, White F (1978) The Indian Ocean Coastal Belt. In: Werger MJA (ed) Biogeography and ecology of southern Africa. Monographiae Biologicae, vol 31. Springer, Dordrecht
Monteiro PMS, van der Plas AK (2006) Low Oxygen Water (LOW) variability in the Benguela system: key processes and forcing scales relevant to forecasting. In: Shannon V, Hempel G, Malanotte-Rizzoli P, Moloney C, Woods J (eds) Benguela: predicting a large marine ecosystem. Large marine ecosystems, vol 14. Elsevier, Amsterdam
Monteiro PMS, van der Plas AK, Bailey GW, Malanotte-Rizzoli P, Duncombe Rae CM, Byrnes D, Pitcher G, Florenchie P, Penven P, Fitzpatrick J, Lass HU (2006) Low Oxygen Water (LOW) forcing scales amenable to forecasting in the Benguela ecosystem. In: Shannon V, Hempel G, Malanotte-Rizzoli P, Moloney C, Woods J (eds) Benguela: predicting a large marine ecosystem. Large marine ecosystems, vol 14. Elsevier, Amsterdam
Morgan JA, LeCain DR, Pendall E, et al. (2011) C4 grasses prosper as carbon dioxide eliminates desiccation in warmed semi-arid grassland. Nature 476:202–205 https://doi.org/10.1038/nature10274
Moriarty R, Buitenhuis ET, Le Quéré C, Gosselin MP (2013) Distribution of known macrozooplankton abundance and biomass in the global ocean. Earth Syst Sci Data 5:241–257
Mucina L, Rutherford MC (2006) The vegetation of South Africa, Lesotho and Swaziland, vol 19. South African National Biodiversity Institute (SANBI), Pretoria
Mudombi S, Nyambane A, von Maltitz GP, Gasparatos A, Johnson FX, Chenene ML, Attanassov B (2018) User perceptions about the adoption and use of ethanol fuel and cookstoves in Maputo, Mozambique. Energy Sustain Dev 44:97–108
Mwaala D (2022) Spatial, seasonal, interannual variability and long-term trends in the diet of Cape fur seals along the Namibian coast (1994–2018). MSc thesis, University of Namibia, Windhoek, Namibia, pp 127
Nakanyala J, Kosmas S, Hipondoka M (2018) The savannas: an integrated synthesis of three major competing paradigms. Int Sci Technol J Namibia 10:108–121
Nelson-Viljoen C, Kyriacou K (2017) Shellfish exploitation strategies at the Pinnacle Point Shell Midden Complex, South Africa, during the later stone age. J Island Coast Archaeol 12:540–557
Nielsen M, Bunkenborg M (2020) Natural resource extraction in the interior: scouts, spirits and Chinese loggers in the forests of northern Mozambique. J South Afr Stud 46:417–433
O'Connor T, Bredenkamp G (2003) Grassland. In: Cowling RM, Richardson DM, Pierce SM (eds) Vegetation of southern Africa. Cambridge University Press, Cambridge
Oesterwind D, Barrett CJ, Sell AF, Núñez-Riboni I, Kloppmann M, Piatkowski U, Wieland K, Laptikhovsky V (2022) Climate change-related changes in cephalopod biodiversity on the North East Atlantic Shelf. Biodivers Conserv 31:1491–1518
O'Hara CC, Villaseñor-Derbez JC, Ralph GM, Halpern BS (2019) Mapping status and conservation of global at-risk marine biodiversity. Conserv Lett 12:e12651
Oliver MJ, Irwin AJ (2008) Objective global ocean biogeographic provinces. Geophys Res Lett 35:L15601
Olson DM, Dinerstein E, Wikramanayake ED, Burgess ND, Powell GVN, Underwood EC, D'amico JA, Itoua I, Strand HE, Morrison JC, Loucks CJ, Allnutt TF, Ricketts TH, Kura Y, Lamoreux JF, Wettengel WW, Hedao P, Kassem KR (2001) Terrestrial ecoregions of the world: a new map of life on earth: a new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. Bioscience 51:933–938
Opdal AF, Brodeur RD, Cieciel K, Daskalov GM, Mihneva V, Ruzicka JJ, Verheye HM, Aksnes DL (2019) Unclear associations between small pelagic fish and jellyfish in several major marine ecosystems. Sci Rep 9:2997
Ortega-Cisneros K, Yokwana S, Sauer W, Cochrane K, Cockcroft A, James N, Potts W, Singh L, Smale M, Wood A, Pecl G (2018a) Assessment of the likely sensitivity to climate change for the key marine species in the southern Benguela system. Afr J Mar Sci 40:279–292
Ortega-Cisneros K, Cochrane KL, Fulton EA, Gorton R, Popova E (2018b) Evaluating the effects of climate change in the southern Benguela upwelling system using the Atlantis modelling framework. Fish Oceanogr 27:489–503
Palmer A, Hoffman M (2003) Nama-Karoo. In: Cowling RM, Richardson DM, Pierce SM (eds) Vegetation of southern Africa. Cambridge University Press, Cambridge
Parr C, Brockett B (1999) Patch-mosaic burning: a new paradigm for savanna fire management in protected areas? Koedoe: African Protected Area Conservation and Science 42(2):117–130. https://doi.org/10.4102/koedoe.v42i2.237
Pillar S (1986) Temporal and spatial variations in copepod and euphausiid biomass off the southern and south-western coasts of South Africa in 1977/78. S Afr J Mar Sci 4:219–229
Postel L, da Silva AJ, Mohrholz V, Lass H-U (2007) Zooplankton biomass variability off Angola and Namibia investigated by a lowered ADCP and net sampling. J Mar Syst 68:143–166
Potts WM, Götz A, James N (2015) Review of the projected impacts of climate change on coastal fishes in southern Africa. Rev Fish Biol Fish 25:603–630
Procheş Ş, Cowling R (2006) Insect diversity in Cape Fynbos and neighbouring South African vegetation. Glob Ecol Biogeogr 15:445–451
Ragoasha N, Herbette S, Cambon G, Veitch J, Reason C, Roy C (2019) Lagrangian pathways in the southern Benguela upwelling system. J Mar Syst 195:50–66
Rebelo AG, Boucher C, Helme N, Mucina L, Rutherford MC (2006) Fynbos Biome. In: Mucina L, Rutherford MC (eds) The vegetation of South Africa, Lesotho and Swaziland. South African National Biodiversity Institute (SANBI), Pretoria
Richardson AJ (2008) In hot water: zooplankton and climate change. ICES J Mar Sci 65:279–295
Richardson DM, van Wilgen BW, Higgins SI, Trinder-Smith TH, Cowling RM, McKell DH (1996) Current and future threats to plant biodiversity on the Cape Peninsula, South Africa. Biodivers Conserv 5:607–647
Riebesell U, Schulz KG, Bellerby RGJ, Botros M, Fritsche P, Meyerhofer M, Neill C, Nondal G, Oschlies A, Wohlers J, Zollner E (2007) Enhanced biological carbon consumption in a high CO2 ocean. Nature 450:545–548
Rittinghaus H (2021) On the distribution, population structure and ecological role of key species of the copepod family Calanidae in the Benguela upwelling system. BSc thesis, University of Bremen, Bremen, Germany, pp 67
Rixen T, Gaye B, Emeis K-C, Ramaswamy V (2019) The ballast effect of lithogenic matter and its influences on the carbon fluxes in the Indian Ocean. Biogeosciences 16:485–503
Rixen T, Lahajnar N, Lamont T, Koppelmann R, Martin B, van Beusekom JEE, Siddiqui C, Pillay K, Meiritz L (2021) Oxygen and nutrient trapping in the southern Benguela upwelling system. Front Mar Sci 8:1367
Robinson WM, Butterworth DS, Plagányi ÉE (2015) Quantifying the projected impact of the South African sardine fishery on the Robben Island penguin colony. ICES J Mar Sci 72:1822–1833
Robison BH (2004) Deep pelagic biology. J Exp Mar Biol Ecol 300:253–272
Robison BH (2009) Conservation of deep pelagic biodiversity. Conserv Biol 23:847–858
Roobaert A, Laruelle GG, Landschützer P, Gruber N, Chou L, Regnier P (2019) The spatiotemporal dynamics of the sources and sinks of CO2 in the global coastal ocean. Glob Biogeochem Cycles 33:1693–1714
Rosa R, Dierssen HM, Gonzalez L, Seibel BA (2008) Large-scale diversity patterns of cephalopods in the Atlantic open ocean and deep sea. Ecology 89:3449–3461
Rosa R, Pissarra V, Borges FO, Xavier J, Gleadall IG, Golikov A, Bello G, Morais L, Lishchenko F, Roura Á (2019) Global patterns of species richness in coastal cephalopods. Front Mar Sci 6:469
Roux J-P, van der Lingen CD, Gibbons MJ, Moroff NE, Shannon LJ, Smith AD, Cury PM (2013) Jellyfication of marine ecosystems as a likely consequence of overfishing small pelagic fishes: lessons from the Benguela. Bull Mar Sci 89:249–284
Rutherford MC, Westfall RH (1986) Biomes of southern Africa - an objective categorization. Memoirs of the Botanical Survey of South Africa, No. 54. National Botanical Institute
Rutherford MC, Westfall RH (1994) Biomes of southern Africa - an objective categorization. Memoirs of the Botanical Survey of South Africa, No. 63. National Botanical Institute
Rutherford MC, Mucina L, Powrie LW (2006) Biomes and bioregions of southern Africa. In: Mucina L, Rutherford MC (eds) The vegetation of South Africa, Lesotho and Swaziland, vol 19. South African National Biodiversity Institute (SANBI), Pretoria
Rykaczewski RR, Dunne JP, Sydeman WJ, García-Reyes M, Black BA, Bograd SJ (2015) Poleward displacement of coastal upwelling-favourable winds in the ocean's eastern boundary currents through the 21st century. J Geophys Res Letters 42:6424–6431
Salvanes AGV, Christiansen H, Taha Y, Henseler C, Seivåg ML, Kjesbu OS, Folkvord A, Utne-Palm AC, Currie B, Ekau W (2018) Variation in growth, morphology and reproduction of the bearded goby (Sufflogobius bibarbatus) in varying oxygen environments of northern Benguela. J Mar Syst 188:81–97
Sankaran M, Hanan NP, Scholes RJ, Ratnam J, Augustine DJ, Cade BS, Gignoux J, Higgins SI, Le Roux X, Ludwig F, Ardo J, Banyikwa F, Bronn A, Bucini G, Caylor KK, Coughenour MB, Diouf A, Ekaya W, Feral CJ, February EC, Frost PG, Hiernaux P, Hrabar H, Metzger KL, Prins HH, Ringrose S, Sea W, Tews J, Worden J, Zambatis N (2005) Determinants of woody cover in African savannas. Nature 438:846–849
Schlebusch CM, Malmström H, Günther T, Sjödin P, Coutinho A, Edlund H, Munters AR, Vicente M, Steyn M, Soodyall H (2017) Southern African ancient genomes estimate modern human divergence to 350,000 to 260,000 years ago. Science 358:652–655
Scholes R (2003) Savanna. In: Cowling R, Richardson D, Pierce S (eds) Vegetation of southern Africa. Cambridge University Press, Cambridge
Scholes RJ, Archer SR (1997) Tree-grass interactions in savannas. Annu Rev Ecol Syst 28:517–544
Scholes R, Walker B (1993) An African savanna: synthesis of the Nylsvley study. Cambridge University Press, Cambridge
Scholes R, Gureja N, Giannecchinni M, Dovie D, Wilson B, Davidson N, Piggott K, McLoughlin C, Van der Velde K, Freeman A, Bradley S, Smart R, Ndala S (2001) The environment and vegetation of the flux measurement site near Skukuza, Kruger National Park. Koedoe 44:73–83
Scholes R, von Maltitz G, Archibald S, Wessels K, van Zyl T, Swanepoel D, Steenkamp K (2014) Terrestrial ecosystem carbon stocks in South Africa. South African National Carbon Sinks Assessment. Department of Environmental Affairs, Pretoria
Schukat A, Bode M, Auel H, Carballo R, Martin B, Koppelmann R, Hagen W (2013) Pelagic decapods in the northern Benguela upwelling system: distribution, ecophysiology and contribution to active carbon flux. Deep-Sea Res I Oceanogr Res Pap 75:146–156
Schukat A, Auel H, Teuber L, Lahajnar N, Hagen W (2014) Complex trophic interactions of calanoid copepods in the Benguela upwelling system. J Sea Res 85:186–196
Schulze BR (1969) The climate of Gobabeb. Scientific Papers of the Namib Desert Research Station 38:5–12
Schwartzlose R, Alheit J, Bakun A, Baumgartner T, Cloete R, Crawford J, Fletcher W, Green-Ruiz Y, Hagen E, Kawasaki T, Lluch-Belda D, Lluch-Cota S, Maccall A, Matsuura Y, Nevárez-Martínez M, Parrish R, Roy C, Serra R, Shust KV, Ward MN, Zuzunaga JZ (1999) Worldwide large-scale fluctuations of sardine and anchovy populations. S Afr J Mar Sci 21:289–347
Shannon L (2009) Benguela Current. In: Steele JH, Thorpe SA, Turekian KK (eds) Encyclopedia of Ocean Sciences, 2nd edn. Academic Press, London, pp 316–327
Shannon L, Pillar S (1986) The Benguela ecosystem. Part III. Plankton. Oceanogr Mar Biol Annu Rev 24:65–170
Shannon LJ, Moloney CL, Jarre A, Field JG (2003) Trophic flows in the southern Benguela during the 1980s and 1990s. J Mar Syst 39:83–116
Siddiqui C, Rixen R, Lahajnar N, van der Plas AK, Louw DC, Lamont T, Pillay K (2023) Regional and global impact of CO2 uptake in the Benguela Upwelling System through preformed nutrients. Nat. Comm. 14:2582. https://doi.org/10.1038/s41467-023-38208-y
Siegfried L, Schmidt M, Mohrholz V, Pogrzeba H, Nardini P, Böttinger M, Scheuermann G (2019) The tropical-subtropical coupling in the Southeast Atlantic from the perspective of the northern Benguela upwelling system. PLoS One 14:e0210083
Sink K, Attwood C, Lombard A, Grantham H, Leslie R, Samaai T, Kerwath S, Majiedt P, Fairweather T, Hutchings L (2011) Spatial planning to identify focus areas for offshore biodiversity protection in South Africa. Final Report for the Offshore Marine Protected Area Project Cape Town: South African National Biodiversity Institute
Sink K, Van der Bank M, Majiedt P, Harris L, Atkinson L, Kirkman S, Karenyi N (2019) South African National Biodiversity Assessment 2018 Technical Report, Vol 4: Marine Realm. South African National Biodiversity Institute, Pretoria
Skowno AL, Thompson MW, Hiestermann J, Ripley B, West AG, Bond WJ (2017) Woodland expansion in South African grassy biomes based on satellite observations (1990–2013): general patterns and potential drivers. Glob Chang Biol 23:2358–2369
Skowno A, Poole C, Raimondo D, Sink K, Van Deventer H, Van Niekerk L, Harris L, Smith-Adao L, Tolley K, Zengeya T, Foden W, Midgley G, Driver A (2019) National Biodiversity Assessment 2018: the status of South Africa’s ecosystems and biodiversity. Synthesis Report South African National Biodiversity Institute, an entity of the Department of Environment, Forestry and Fisheries, Pretoria
Smit AJ, Bolton JJ, Anderson RJ (2017) Seaweeds in two oceans: beta-diversity. Front Mar Sci 4:404
Smith HE, Jones D, Vollmer F, Baumert S, Ryan CM, Woollen E et al. (2019) Urban energy transitions and rural income generation: sustainable opportunities for rural development through charcoal production. World Dev 113:237–245
Starke C, Ekau W, Moosdorf N (2019) Enhanced productivity and fish abundance at a submarine spring in a coastal lagoon on Tahiti, French Polynesia. Front Mar Sci 6:809. https://doi.org/10.3389/fmars.2019.00809
Staver AC, Archibald S, Levin SA (2011) The global extent and determinants of savanna and forest as alternative biome states. Science 334:230–232
Stevens N, Erasmus BFN, Archibald S, Bond WJ (2016) Woody encroachment over 70 years in South African savannahs: overgrazing, global change or extinction aftershock? Philos Trans R Soc B: Biol Sci 371:20150437
Stramma L, Johnson GC, Sprintall J, Mohrholz V (2008) Expanding oxygen-minimum zones in the tropical oceans. Science 320:655–658
Stuut J-BW, Crosta X, Van der Borg K, Schneider R (2004) Relationship between Antarctic sea ice and southwest African climate during the late quaternary. Geology 32:909–912
Sutton TT, Clark MR, Dunn DC, Halpin PN, Rogers AD, Guinotte J, Bograd SJ, Angel MV, Perez JAA, Wishner K, Haedrich RL, Lindsay DJ, Drazen JC, Vereshchaka A, Piatkowski U, Morato T, Blachowiak-Samolyk K, Robison BH, Gjerde KM, Pierrot-Bults A, Bernal P, Reygondeau G, Heino M (2017) A global biogeographic classification of the mesopelagic zone. Deep-Sea Res Pt I 126:85–102
Sweijd N, Smit A (2020) Trends in sea surface temperature and chlorophyll-a in the seven African Large Marine Ecosystems. Environ Dev 36:100585
Sydeman WJ, García-Reyes M, Schoeman DS, Rykaczewski RR, Thompson SA, Black BA, Bograd SJ (2014) Climate change and wind intensification in coastal upwelling ecosystems. Science 345:77–80
Teske PR, Von der Heyden S, McQuaid CD, Barker NP (2011) A review of marine phylogeography in southern Africa. S Afr J Sci 107:1–11
Teuber L, Schukat A, Hagen W, Auel H (2013) Distribution and ecophysiology of calanoid copepods in relation to the oxygen minimum zone in the eastern tropical Atlantic. PLoS One 8:e77590
Teuber L, Hagen W, Bode M, Auel H (2019) Who is who in the tropical Atlantic? Functional traits, ecophysiological adaptations and life strategies in tropical calanoid copepods. Prog Oceanogr 171:128–135
Tolley KA, Chase BM, Forest F (2008) Speciation and radiations track climate transitions since the Miocene climatic optimum: a case study of southern African chameleons. J Biogeogr 35:1402–1414
Tolley K, da Silva J, Jansen van Vuuren B (2019) South African National Biodiversity Assessment 2018 Technical Report Vol 7: Genetic Diversity. South African National Biodiversity Institute, Pretoria
Tyler T (2016) Examining the feeding ecology of two mesopelagic fishes (Lampanyctodes hectoris & Maurolicus walvisensis) off the West Coast of South Africa using stable isotope and stomach content analyses. MSc thesis, University of Cape Town, 67 pp
Utne-Palm AC, Salvanes AG, Currie B, Kaartvedt S, Nilsson GE, Braithwaite VA, Stecyk JA, Hundt M, van der Bank M, Flynn B (2010) Trophic structure and community stability in an overfished ecosystem. Science 329:333–336
van der Lingen CD (2021) Adapting to climate change in the South African small pelagic fishery. In: Bahri T, Vasconcellos M, Welch DJ, Johnson J, Perry RI, Ma X, Sharma R (eds) Adaptive management of fisheries in response to climate change. FAO Fisheries and Aquaculture Technical Paper No 667. FAO, Rome
van der Lingen CD, Hampton I (2018) Climate change impacts, vulnerabilities and adaptations: Southeast Atlantic and Southwest Indian Ocean marine fisheries. In: Barange M, Bahri T, Beveridge MCM, Cochrane KL, Funge-Smith S, Poulain F (eds) Impacts of climate change on fisheries and aquaculture: synthesis of current knowledge, adaptation and mitigation options, FAO Fisheries and Aquaculture Technical Paper No 627. FAO, Rome
van der Lingen CD, Miller TW (2014) Spatial, ontogenetic and interspecific variability in stable isotope ratios of nitrogen and carbon of Merluccius capensis and Merluccius paradoxus off South Africa. J Fish Biol 85:456–472
van der Lingen CD, Fréon P, Hutchings L, Roy C, Bailey GW, Bartholomae C, Cockcroft AC, Field JG, Peard KR, van der Plas AK (2006a) Forecasting shelf processes of relevance to living marine resources in the BCLME. In: Shannon V, Hempel G, Malanotte-Rizzoli P, Moloney C, Woods J (eds) Large Marine Ecosystems. Elsevier, Amsterdam
van der Lingen CD, Hutchings L, Field JG (2006b) Comparative trophodynamics of anchovy Engraulis encrasicolus and sardine Sardinops sagax in the southern Benguela: are species alternations between small pelagic fish trophodynamically mediated? S Afr J Mar Sci 28:465–477
van der Lingen CD, Shannon LJ, Cury P, Kreiner A, Moloney CL, Roux J-P, Vaz-Velho F (2006c) Resource and ecosystem variability, including regime shifts, in the Benguela Current System. In: Shannon V, Hempel G, Malanotte-Rizzoli P, Moloney C, Woods J (eds) Benguela: predicting a Large Marine Ecosystem, vol 14. Elsevier, Amsterdam
van der Lingen CD, Hutchings L, Lamont T, Pitcher GC (2016) Climate change, dinoflagellate blooms and sardine in the southern Benguela Current Large Marine Ecosystem. Environ Dev 17:230–243
van der Sleen P, Zuidema PA, Morrongiello J, Ong JLJ, Rykaczewski RR, Sydeman WJ, Di Lorenzo E, Black BA (2022) Interannual temperature variability is a principal driver of low-frequency fluctuations in marine fish populations. Commun Biol 5:1–8
van Rensburg B, Chown S, Gaston K (2002) Species richness, environmental correlates, and spatial scale: a test using South African birds. Am Nat 159:566–577
van Waarden C (2002) The early iron age of Botswana. In: Oxford Research Encyclopedia of Anthropology. Oxford University Press, Oxford
van Wilgen B, Richardson D, Seydack A (1994) Managing Fynbos for biodiversity: constraints and options in a fire-prone environment. S Afr J Sci 90:322–329
van Wilgen BW, Forsyth GG, De Klerk H, Das S, Khuluse S, Schmitz P (2010) Fire management in Mediterranean-climate shrublands: a case study from the Cape Fynbos, South Africa. J Appl Ecol 47:631–638
Vansina J (1994) A slow revolution: farming in subequatorial Africa. Azania: Archaeol Res Africa 29:15–26
Verheye H, Hagen W, Auel H, Ekau W, Loick N, Rheenen I, Wencke P, Jones S (2005) Life strategies, energetics and growth characteristics of Calanoides carinatus (Copepoda) in the Angola-Benguela frontal region. Afr J Mar Sci 27:641–651
Verheye H, Hutchings L, Huggett J, Painting S (1992) Mesozooplankton dynamics in the Benguela ecosystem, with emphasis on the herbivorous copepods. In: Payne A, Brink K, Mann K, Hilborn R (eds) Benguela Trophic Functioning. S Afr J Mar Sci, vol 12. pp 561–584
Verheye H, Hutchings L, Peterson W (1991) Life history and population maintenance strategies of Calanoides carinatus (Copepoda: Calanoida) in the southern Benguela ecosystem. S Afr J Mar Sci 11:179–191
Verheye HM, Hutchings L, Huggett JA, Carter RA, Peterson WT, Painting SJ (1994) Community structure, distribution and trophic ecology of zooplankton on the Agulhas Bank with special reference to copepods. S Afr J Sci 90:154–165
Verheye HM, Lamont T, Huggett JA, Kreiner A, Hampton I (2016) Plankton productivity of the Benguela Current Large Marine Ecosystem (BCLME). Environ Dev 17:75–92
Verheye HM, Rogers C, Maritz B, Hashoongo V, Arendse LM, Gianakouras D, Giddey CJ, Herbert V, Jones S, Kemp AD, Ruby C (2001) Variability of zooplankton in the region of the Angola-Benguela front during winter 1999: BENEFIT Marine Science. S Afr J Sci 97:257–258
von Maltitz G, Knowles T, Wiese L, Scholes R, Pienaar M, Davies C, Steenkamp K (2020) National Terrestrial Carbon Sinks Assessment: Technical Report (2020). Technical Report CSIR. Department of Environment, Forestry and Fisheries, Pretoria, South Africa
von Maltitz GP, Lindeque GHL, Kellner K (2018) A changing narrative on desertification and degradation in South Africa. Desertification: past, current and future trends. Nova Science Publishers, Inc, New York
von Maltitz GP, Mucina L, Geldenhuys C, Lawes M, Eeley H, Adie H, Vink D, Fleming G, Bailey C (2003) Classification system for South African indigenous forests: an objective classification for the Department of Water Affairs and Forestry. Environmental Report ENV-P-C 2003-017. CSIR, Pretoria, South Africa
Vorrath M-E, Lahajnar N, Fischer G, Libuku VM, Schmidt M, Emeis K-C (2018) Spatiotemporal variation of vertical particle fluxes and modelled chlorophyll a standing stocks in the Benguela upwelling system. J Mar Syst 180:59–75
Wang D, Gouhier TC, Menge BA, Ganguly AR (2015) Intensification and spatial homogenization of coastal upwelling under climate change. Nature 518:390–394
Werner T, Buchholz F (2013) Diel vertical migration behaviour in euphausiids of the northern Benguela Current: seasonal adaptations to food availability and strong gradients of temperature and oxygen. J Plankton Res 35:792–812
White F (1983) The vegetation of Africa, a descriptive memoir to accompany the UNESCO/AETFAT/UNSO vegetation map of Africa. Nat Resour Res 20:1–356
Wilhelm MR, Kirchner CH, Roux JP, Jarre A, Iitembu JA, Kathena JN, Kainge P (2015) Biology and fisheries of the shallow-water hake (Merluccius capensis) and the deep-water hake (Merluccius paradoxus) in Namibia. In: Arancibia H (ed) Hakes: biology and exploitation. Wiley Blackwell, Oxford
Wilhelm MR, Black BA, Lamont T, Paulus SC, Bartholomae C, Louw DC (2020) Northern Benguela Merluccius paradoxus annual growth from otolith chronologies used for age verification and as indicators of fisheries-induced and environmental changes. Front Mar Sci 7:315
Woodd-Walker RS (2001) Spatial distributions of copepod genera along the Atlantic meridional transect. Hydrobiologia 453:161–170
Woodd-Walker RS, Ward P, Clarke A (2002) Large-scale patterns in diversity and community structure of surface water copepods from the Atlantic Ocean. Mar Ecol Prog Ser 236:189–203
Woollen E, Ryan CM, Baumert S, Vollmer F, Grundy I, Fisher J, Fernando J, Luz A, Ribeiro N, Lisboa SN (2016) Charcoal production in the mopane woodlands of Mozambique: what are the trade-offs with other ecosystem services? Philos Trans R Soc B: Biol Sci 371:20150315
Wren CD, Botha S, De Vynck J, Janssen MA, Hill K, Shook E, Harris JA, Wood BM, Venter J, Cowling R (2020) The foraging potential of the Holocene Cape South Coast of South Africa without the Palaeo-Agulhas plain. Quat Sci Rev 235:105789
Acknowledgments
We appreciate the valuable comments of an anonymous reviewer and the editor, Christian Brümmer, on a previous version of this chapter. We thank Solvin Zankl for taking and providing the photographs of marine organism for Figs. 2.4 and 2.5 (www.solvinzankl.com). The SPACES II projects TRAFFIC, CASISAC and BANINO were funded by the German Federal Ministry of Education and Research (BMBF), FKZ 03F0797, 03F0796 and 03F0795.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2024 The Author(s)
About this chapter

Cite this chapter
Sell, A.F. et al. (2024). Unique Southern African Terrestrial and Oceanic Biomes and Their Relation to Steep Environmental Gradients. In: von Maltitz, G.P., et al. Sustainability of Southern African Ecosystems under Global Change. Ecological Studies, vol 248. Springer, Cham. https://doi.org/10.1007/978-3-031-10948-5_2
Download citation
DOI: https://doi.org/10.1007/978-3-031-10948-5_2
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-10947-8
Online ISBN: 978-3-031-10948-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)